|
| |
| |
| |
6. Vliegende stukjes boomschors
De camouflage van de heidevlinder
Een van de meest algemeen voorkomende vlindersoorten van de Hulshorster stuifzanden en desalniettemin een van de minst opvallende was de heidevlinder Eumenis (= Satyrus) semele. Toen ik dit insekt voor de eerste maal zag, maakte het diepe indruk op mij vanwege zijn vrijwel volmaakte camouflage. Wij stonden te kijken naar de verschillende insekten die bijeen zaten op een ‘bloedende’ berk. Talrijke berken en eiken in dit gebied waren aangetast door de wilgehoutrups, die zich in het levende hout boort. Via de wonden die deze grote rupsen maakten, vloeide sap naar buiten en de doordringende geur hiervan lokte talrijke gastronomisch ingestelde insekten aan, zoals atalanta's, rouwmantels, kleine vossen, dagpauwogen, rode bosmieren, allerlei soorten vliegen, diverse wespen en kevers. Andere bezoekers hadden geen belangstelling voor de berkewijn zelf, maar meer voor de zich hieraan te goed doende gasten. Zwermen wespen (Vespa media en Vespa crabro) kwamen hier vliegen en zelfs grote vlinders vangen; op vliegen jagende graafwespen (Mellinus) vingen niet alleen hier hun vliegen, maar groeven hun nesten aan de voet van dergelijke bomen, om voedsel bij de hand te hebben; ze bouwden hun holletjes als het ware bij de supermarkt.
Terwijl wij zagen hoe een rouwmantel de boom naderde, wat hij zoals gewoonlijk tegen de wind in deed, ongetwijfeld aangelokt door de geur, maakte zich een klein stukje schors los van de boom, schoot door de lucht op de vlinder af, fladderde er enkele seconden omheen, liet hem dan weer met rust en liet zich op de grond vallen, waar het even plotseling verdween als het verschenen was.
Toen ik voorzichtig de plek naderde waar ik het vreemde ding de grond had zien raken, zag ik niets totdat een klein stukje vuil zich plotseling in de lucht verhief en rakelings achter mij langs schoot, terug naar de boom, waar het onmiddellijk uit het gezicht verdween zodra het de schors had bereikt. Dit was een heidevlinder of, zoals wij hem noemden, een stukje boombast met vleugels.
De camouflage van dit insekt was werkelijk verbijsterend. Als het dier in rust zat, hield hij zoals alle vlinders zijn vleugels opgevouwen: de voorvleugels omlaaggeklapt tussen de achtervleugels, zodat
| |
| |
| |
| |
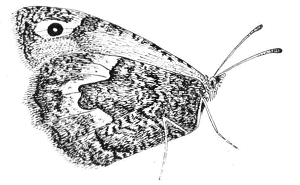
Een heidevlinder vertoont zijn opvallende oogvlekken.
alleen de onderkant van de achtervleugels zichtbaar was. Als men verder dan enkele meters van het dier vandaan was, had deze onderzijde een onopvallende grijze kleur, en was zij zodanig gespikkeld en gestreept dat het patroon leek op het onregelmatige oppervlak van een boombast. Wanneer men echter deze vleugels onder een loep bekeek, ontdekte men dat het schijnbaar kleurloze patroon samengesteld was uit schubben die op zichzelf schitterende kleuren hadden, variërend van zeer donkerbruin tot wit of goudgeel. Zij waren echter zodanig gerangschikt dat er geen grote vlakken van een bepaalde kleur te zien waren, zodat de algehele indruk die van een grijs oppervlak was. Deze zelfde kleuren vond men terug aan de bovenzijde van de achtervleugels en aan beide zijden van de voorvleugels, maar daar waren zij wel gerangschikt in grote, meer uniforme kleurvelden. Deze bovenzijden waren donkerbruin, met brede okerkleurige banden waarin zwarte ronde vlekken met in het midden een witte stip voorkwamen, twee op iedere vleugel. Dit patroon maakte de bovenzijde van de vleugels zeer kleurrijk en opvallend. De heldere kleuren waren zichtbaar tijdens de vlucht, maar verdwenen onmiddellijk zodra het dier zich ergens neerzette. Het camouflerende effect en de middelen waarmee dit bereikt werd, waren de moeite van een nauwkeurig onderzoek waard. Het boomschors-patroon was niet beperkt tot de onderzijde van de achtervleugels, maar kwam ook voor op de allerbuitenste uiteinden van de voorvleugels, die zichtbaar bleven als de vleugels in rust samengevouwen waren; de grens viel precies samen met de rand van de achtervleugels; geen millimetertje kleur was zichtbaar als de vlinder stil zat. Evenmin trof men het
schorsige patroon aan op vleugeldelen
| |
| |
die in rust onzichtbaar waren. De bewegingen van de heidevlinder vertoonden dezelfde specialisaties die men ook bij talrijke andere gecamoufleerde dieren ziet: ofwel zij vlogen, en dan zeer snel, of zij zaten volkomen roerloos. De overgang van een razendsnelle vlucht naar complete bewegingloosheid was zo abrupt, dat wij een heidevlinder altijd onmiddellijk uit het oog verloren als hij ergens landde.
| |
Paargedrag
Hoe fascinerend de heidevlinders ook waren, toch besteedden wij niet zoveel aandacht aan ze omdat wij volledig in beslag genomen werden door onze waarnemingen bij de graafwespen. Maar toen in de late jaren dertig de aantallen van Philanthus begonnen te verminderen, en verdere oriëntatie-experimenten op grote schaal niet langer mogelijk waren, richtten wij onze aandacht op andere insekten en ontdekten wij al gauw dat de heidevlinder uitstekend studiemateriaal vormde. Wij herinnerden ons dat wij hem achter andere vlinders aan hadden zien vliegen en vroegen ons af of dit regelmatig gebeurde en zo ja, waarom.
Een paar dagen van intensief observeren in de buurt van een bloedende boom gaf ons het antwoord hierop en maakte ons duidelijk dat hier veelbelovende mogelijkheden voor veldonderzoek lagen. Wij ontdekten dat op zonnige dagen de mannetjes-heidevlinders (die kleiner waren dan de wijfjes, een lichter gekleurde band op hun gecamoufleerde vleugels bezaten en bij wie de bovenzijde van de vleugels lichter bruin waren) bepaalde uitkijkposten betrokken op bloedende bomen of ergens anders, maar steeds op of een weinig boven de grond. Gewoonlijk zaten zij volkomen bewegingloos, maar als er een andere vlinder over kwam vliegen, schoten zij daar onveranderlijk onmiddellijk achteraan. Dikwijls keerden zij plotseling terug als ze de vlinder bijna ingehaald hadden, maar als het een vrouwelijke heidevlinder betrof, werd het een wilde jacht. Sommige van deze wijfjes vlogen er zo snel als zij konden vandoor. Anderen zetten zich ergens neer en dan streken ook de achtervolgende mannetjes neer en liepen vervolgens naar het wijfje toe. Daarop liep het wijfje ofwel weg, met heftig vleugelgeklapper, ofwel zij bleef roerloos zitten. Een wijfje dat met haar vleugels sloeg werd door het mannetje verder met rust gelaten, maar wanneer er een ging zitten en haar vleugels samenvouwde, voerde het mannetje een fascinerende, uiterst ingewikkelde show op. Nadat hij vlak bij het wijfje
| |
| |
was neergestreken, liep hij om haar heen, totdat hij haar recht in het gezicht keek. Dan hief hij met eigenaardige korte rukjes zijn voorvleugels wat op, die daarbij voortdurend trilden en tenslotte sloeg hij het wijfje, tijdens een laatste voorwaartse rukbeweging, met zijn vleugels. Al die tijd hield hij zijn vleugels geheel of gedeeltelijk opgevouwen. Vervolgens trok hij zijn voorvleugels iets terug en begon de voorste randen ervan in een snelle opeenvolging van bewegingen beurtelings te openen en te sluiten, hetgeen een zeer eigenaardige beweging was, aangezien de vleugels voor het overige samengevouwen bleven.
Terwijl hij hiermee bezig was, hield hij zijn antennen zodanig dat ze een rechte hoek maakten met zijn lichaam, terwijl hij ze ritmisch op en neer bewoog zodat de uiteinden een halve of hele cirkel per seconde beschreven. Het wijfje strekte haar antennen uit naar de vleugels van het mannetje. Na enkele seconden zo met zijn vleugels gewaaierd te hebben, vouwde het mannetje plotseling zijn vleugels helemaal open, bewoog zich dan langzaam naar voren en, de antennen van het wijfje tussen zijn vleugels nemend, drukte hij deze laatste langzaam maar krachtig samen. Hierna liep hij weer om haar heen totdat hij enigszins schuin achter haar stond en terwijl hij zijn achterlijf zijwaarts kromde, maakte hij contact met haar genitaliën en vond de paring plaats.
Dit was het hele paringsproces, maar wij hebben het maar enkele malen zo volledig in de natuur waargenomen. De meeste mannetjes vingen bot bij de wijfjes; zoals wij later ontdekten werd dit veroorzaakt door het feit dat de meeste, zo niet alle wijfjes slechts eenmaal paarden en de mannetjes van veraf geen onderscheid konden maken tussen maagdelijke wijfjes en wijfjes die al gepaard hadden. Zij vlogen achter alle wijfjes aan en ontdekten pas later of een wijfje al dan niet op hun avances inging.
Het was dus duidelijk dat het plotseling achtervolgen van hetzij een wijfje, hetzij een vreemde vlindersoort de inleiding van zijn baltsgedrag vormde. Het leek erop dat het zien van onverschillig wat voor vlinder de mannetjes stimuleerde tot deze seksuele achtervolging en alsof prikkels, die hij ontving nadat hij het andere insekt genaderd was, hem in staat stelden te zien of het een wijfje van zijn eigen soort was of een ander insekt. Het hele baltsritueel leek uit een keten van afzonderlijke reacties te bestaan. In die periode waren wij vooral geïnteresseerd in het onderzoek van prikkelsituaties die enkelvoudige reacties opwekten en hier leek zich een uitstekend object voor een dergelijk onderzoek aan te dienen.
| |
| |

Observatie van een rouwmantel, die juist uit de pop komt.
| |
| |
| |
Op de vlinderjacht
Wij, dat wil zeggen B.J.D. Meeuse, L.K. Boerema, W.W. Varossieau zaliger en ikzelf, startten ons onderzoek met het wat nader bestuderen van de seksuele achtervolging. Het was ons al snel duidelijk dat dit een reactie vormde op een visuele prikkelsituatie van een zeer onspecifieke aard, want de mannetjes gingen achter de meest vreemde voorwerpen aan. Wij zagen hen 25 verschillende Lepidoptera-soorten achtervolgen, waaronder zulke uiteenlopende vormen als rouwmantels, heideblauwtjes en hagehelden. Ook talrijke andere insekten werden achternagezeten: mestkevers, wespen, verschillende soorten libellen, sprinkhanen zoals Stenobothrus en Oedipoda, en zelfs vogels als koolmezen, boekvinken en grote lijsters. Deze waarnemingen duidden er al op dat de effectieve stimuli van visuele aard moesten zijn, en dit vermoeden werd nog gestaafd door tamelijk grappige situaties waarin de mannetjes achter hun eigen schaduw aanvlogen.
Wij besloten om hier nader op in te gaan en de natuurlijke experimenten die wij hadden waargenomen, uit te breiden door meer systematisch en volgens vooropgezet plan proeven te gaan doen.

| |
| |
Wij vervaardigden papieren modellen van vlinders, bonden deze met dun garen aan stokken van ongeveer een meter lang en vertoonden met behulp van deze ‘vishengels’ de modellen aan de rustende mannetjes, waarbij wij ieder model van zo'n twee meter afstand naar het mannetje toe lieten ‘vliegen’ en op dertig centimeter afstand lieten passeren. De mannetjes reageerden gretig op deze modellen, en zo vormde dit het begin van een onderzoek dat ons vele seizoenen lang tot groot vermaak van onze vrienden bezighield. Met drieën of vieren doorkruisten wij voortdurend vele kilometers heidevlinder-territorium en boden ieder mannetje dat wij ontwaarden op min of meer dezelfde manier onze modellen aan. Het zou het beste zijn geweest als wij een vast aantal proeven hadden gedaan, waarbij wij voor ieder mannetje een constant aantal modellen hadden gebruikt, maar dit was in het veld absoluut onmogelijk, want de mannetjes vlogen steeds na een of enkele proeven weg en wij konden ze niet altijd lang genoeg achtervolgen om alle proeven die wij wilden doen, met hen uit te voeren. Voor zover mogelijk boden wij ieder model driemaal snel achter elkaar aan (met vijf tot tien seconden tussenpauze). Dan wachtten wij ongeveer twintig seconden en boden op dezelfde manier een ander model aan. Wij gingen hiermee net zolang door, steeds een ander model aanbiedend, totdat het mannetje ervandoor ging. De duur van een experiment werd dus bepaald door onze mannetjes; deze varieerde van zes tot enkele honderden malen een model aanbieden. Wij veranderden de volgorde waarin wij de verschillende modellen aanboden naar het ons uitkwam (niet zo verfijnd dat het werkelijk at random gebeurde) en wisselden ook onder elkaar ieder uur onze modellen, om mogelijke
individuele verschillen in ‘hengeltechniek’ uit te schakelen.
In totaal voerden wij zo'n 50.000 experimenten uit. Voor elk model werden de positieve reacties en het aantal malen dat zo'n model aangeboden werd, genoteerd en het aantal positieve reacties per 100 maal aanbieden gebruikten wij als een maat voor de signaalwaarde van een model. Wij vergeleken alleen modellen van een reeks met elkaar, want experimenten die op twee verschillende dagen waren uitgevoerd waren natuurlijk niet met elkaar te vergelijken vanwege weersveranderingen en andere variabelen die wij niet in onze macht hadden.
Wij boden (op zijn zachtst uitgedrukt) een merkwaardige aanblik in onze uitermate praktische maar nauwelijks elegant te noemen veldplunje, bestaande uit een korte broek, een breedgerande strooien hoed en zonnebril, ieder van ons gewapend met twee hengels, voor- | |
| |
zien van dansende papieren vlindermodellen (waarvan sommige felgekleurd), voortdurend proberend om niet met onze hengels in struikgewas of pollen heide te blijven hangen, strak turend naar een van onze heidevlinders terwijl wij deze trachtten te volgen op zijn onberekenbare route, erachteraan rennend, plotseling stilstaand, het dier besluipend, dan zorgvuldig ons hengelritueel herhalend, driemaal maar liefst en tenslotte het een en ander opschrijvend; en dit alles met gespannen, doodernstige gezichten. Geen wonder dat wij nieuwsgierige en verwonderde blikken trokken van een toevallig langskomende boer of toerist die ons zo aan het werk zag.
Onze eerste serie proeven diende om uit te vinden of de mannetjes reageerden op de details van het vrouwelijke kleurpatroon. Wij hadden al gezien dat zij even gretig achter zulke uiteenlopende vlinders als heidevlinders en atalanta's aangingen, maar wat wij wilden weten was, of een afwijkend gekleurd model even effectief was als een normaal Eumenis-wijfje. Bij deze eerste reeks proeven begonnen wij met het aanbieden van drie modellen achter elkaar: een egaal grijsbruin, geknipt uit pakpapier, een met behulp van waterverf vervaardigde, zo exact mogelijke kopie van het echte patroon, en een derde model waaraan wij echte vleugels van een wijfjesvlinder hadden geplakt. Bij een andere reeks experimenten voegden wij hier nog een egaal gekleurd model aan toe, met dezelfde aanplak-vleugels. Met deze modellen deden wij meer dan 2000 proeven en noteerden meer dan 1500 positieve reacties. Geen enkel model wekte meer reacties op dan ieder ander en het was ons dus duidelijk dat de details van het kleurpatroon volslagen onbelangrijk waren voor het mannetje, althans wat betreft het opwekken van de achtervolgingsreactie. Feitelijk hadden wij niet veel anders verwacht, want de scherpte van de Eumenis-ogen is waarschijnlijk zodanig dat, onder de omstandigheden waarbij wij onze experimenten uitvoerden, de mannetjes onmogelijk details konden waarnemen. Desalniettemin voerden wij deze experimenten toch uit om er zeker van te kunnen zijn dat het eenvoudige papieren model te gebruiken was als een nagenoeg optimaal standaardmodel, waarmee wij alle andere modellen zouden kunnen vergelijken.
Onze volgende stap was het testen van het effect van kleur door identieke modellen van vijf verschillende kleuren aan te bieden: rood, geel, groen, blauw en bruin (de standaardkleur), maar ook wit en zwart. Alle papieren modellen waren geknipt van het bekende, uit Duitsland afkomstige Hering-papier, dat een standaardkleur heeft en waarvan wij de spectrale reflectie hadden gemeten. In to- | |
| |
taal noteerden wij gedurende twaalf reeksen experimenten meer dan 6000 reacties en tot onze verbazing bestond er geen merkbaar verschil tussen de kleuren. De standaardkleur was even effectief als de rode tint en verschilde nauwelijks van zwart, groen, blauw en geel. Alleen de witte modellen waren duidelijk minder effectief. Zo er al iets uit deze experimenten bleek, was het dat de zwarte papiertjes de meeste reacties leken uit te lokken, maar dat was dan ook alles. Wij gingen dit nog eens na tijdens een aparte reeks proeven waarin wij zwarte en witte modellen vergeleken met twee tinten grijs. De meer dan 1000 reacties die wij bij dit experiment verzamelden, waren zodanig verdeeld dat het zwarte model de hoogste score behaalde, nl. 52, grijs nr. 15 (de donkerste tint grijs) een score van 50, grijs nr. 7 (de lichte tint grijs) 47 en wit 42.
Voor zover het om kleur en schaduwing ging, mocht men hieruit dus concluderen dat een mannetje vooral gestimuleerd werd door een donker wijfje, van welke kleur dan ook. De experimenten met de gekleurde modellen lieten zelfs vermoeden dat een zwart model nog meer voldeed dan een model met de normale kleur.
Nu leek dit nogal tegenstrijdig. De mannetjes vertoonden geen voorkeur voor een speciale kleur en gedroegen zich alsof zij kleurenblind waren. En toch hadden wij dikwijls heidevlinders zich aan bloemen zien voeden en waren wij ervan overtuigd dat zij wel degelijk in staat waren om verschillende kleuren van elkaar te onderscheiden, aangezien zij een voorkeur hadden voor bloemen van allerlei schakeringen blauw.
| |
Kleuren-zien
Wij besloten om het voedingsgedrag wat nader te bestuderen. Voor dit doel bouwden wij een grote (5×5×2 meter) gazen kooi waarin wij de vlinders konden laten rondvliegen en bloemmodellen laten bezoeken als wij dat wilden. Wij legden, in de vorm van een schaakbord, rechthoekige stukken papier uit, die alle kleuren van de regenboog, van rood tot violet, en alle dertig kleurschakeringen van de standaard-Heringreeks vertoonden. Vervolgens vingen wij een dozijn heidevlinders, lieten deze vrij in de kooi en wachtten op de dingen die komen zouden, terwijl wij ons op ons gemak en onopvallend in een hoek van de kooi posteerden. De zon brandde, het witte gaas was ongelooflijk schel aan onze ogen, de atmosfeer zeer stoffig en de observeerders voelden zich uitermate slaperig. Het enthousi- | |
| |
asme kelderde tot een voordien ongekend minimum als een hele dag wachten geen enkele reactie op ook maar één van de papiertjes opleverde. Aanvankelijk dachten wij dat de vlinders geen honger hadden of van slag waren door de ongewone omgeving. Maar toen wij hen echte bloemen aanboden, gingen de meeste van hen zich daaraan te goed doen.
Wij veronderstelden toen dat zij misschien toch eerder reageerden op de geur van de bloemen dan op hun kleur. In elk geval werden hun bezoeken aan de bloedende bomen volledig door de geur bepaald. Om dit na te gaan namen wij nu een paar lapjes, besprenkelden die met verschillende parfums (rozen, lavendel, seringen) en legden ze in de kooi. De vlinders reageerden hier inderdaad op, maar op een volkomen onverwachte wijze. Zij liepen rond, opgewonden met hun antennen trommelend, maar gingen niet op de lapjes af. In plaats daarvan zag Meeuse plotseling een van de vlinders omlaag komen vliegen en landen op het blauwe overhemd dat hij droeg. Hij begreep onmiddellijk wat dit betekende en sloeg zichzelf voor het hoofd dat hij daar niet eerder aan gedacht had. Een Duitse onderzoeker, Dora Ilse, had al eens geschreven dat de geur van bloemen voor sommige vlinders noodzakelijk is om hun reactie op de kleur op te wekken; klaarblijkelijk behoorde ook Eumenis tot deze vlindersoorten. Deze veronderstelling bleek inderdaad juist te zijn en vanaf dat moment werden de vlinders zowel met geuren als met gekleurde papiertjes geconfronteerd. De observeerders, eveneens hieraan blootgesteld, werden bijna misselijk van de bedwelmende geur in de kooi, maar zij werden na enkele dagen beloond met een prachtige eenduidige grafiek: vrijwel elk van de bijna 100 reacties was gericht op hetzij een blauw, hetzij een geel papiertje. Het betrof hier dus een duidelijk voorbeeld van een verschijnsel waarvan wij nu weten dat het zeer algemeen moet voorkomen: de innerlijke toestand van een dier, zijn ‘gemoedstoestand’ is van grote invloed op de prikkels die het uit zijn omgeving opneemt. Vooral deze proef liet dit duidelijker zien dan gewoonlijk, omdat de papiertjes die wij de
hongerige mannetjes aanboden, volkomen identiek in kleur waren aan de modellen die wij tijdens onze hengel- experimenten aan seksueel actieve mannetjes hadden aangeboden.
Voor dat ogenblik lieten wij de zaak verder rusten; wij stelden alleen vast dat seksueel actieve mannetjes op veel minder prikkels reageerden dan hun ogen in staat waren te ontvangen. Men weet tegenwoordig dat dit een zeer algemeen voorkomend verschijnsel
| |
| |
is. Het vormt een interessant fysiologisch probleem, want het bete kent dat, op de een of andere manier, opgenomen informatie die wel de zintuigen passeert, uiteindelijk toch niet terechtkomt bij de motorische centra die verantwoordelijk zijn voor de spierbewegingen; en of deze blokkering al dan niet plaatsvindt, wordt bepaald door de innerlijke toestand van het dier. Aan dit probleem werkt men nu in verscheidene laboratoria. Evenals bij verschillende andere vraagstukken is ook hier sprake van een samengaan van gedragsonderzoek in het veld en neurofysiologisch onderzoek in het laboratorium.
| |
Andere variaties op het model
Om terug te keren naar onze hengel-experimenten: wij deden vervolgens een reeks proeven waarbij wij modellen van verschillende vorm met elkaar vergeleken: een standaardmodel dat de vorm van een vlinder had, een rond model en drie rechthoeken van verschillende afmetingen (16 × 1; 8 × 2; 5 × 3,2 centimeter). Het oppervlak van ieder model was gelijk. De mannetjes vertoonden op elk van de modellen even duidelijke reacties, behalve op de langste rechthoek, waarvoor zij minder belangstelling hadden. Later ontdekten wij echter dat de fladderende bewegingen van de modellen een belangrijke prikkel vormden en aangezien dit langgerekte model niet zo opvallend fladderde als de rest, is de lage score wellicht eerder het gevolg geweest van de beweging dan van de vorm. In elk geval was het duidelijk dat vormkenmerken geen invloed hadden op het opwekken van de seksuele achtervolging. In hoeverre dit een gevolg was van een onvermogen om onderscheid te maken tussen de diverse vormen, weten wij niet.
Toen wij een zelfde reeks experimenten wilden doen met modellen van verschillende afmeting, raakten wij in moeilijkheden. Allereerst ontdekten wij dat, toen wij onze modellen op de gebruikelijke manier aanboden door ze op ongeveer twee meter afstand in het blikveld van het mannetje te brengen en dan langzaam dichterbij te komen, de effectiviteit van de standaardmodellen groter was dan die van zowel de kleinere als de grotere modellen. Sterker nog: wij stelden vast dat de grotere modellen de mannetjes dikwijls wegjoegen in plaats van ze aan te trekken. Vooral op korte afstand was dit schrikeffect zeer duidelijk aanwezig. Daarom veranderden wij van strategie en maakten wij modellen van verschillende grootte, die op
| |
| |
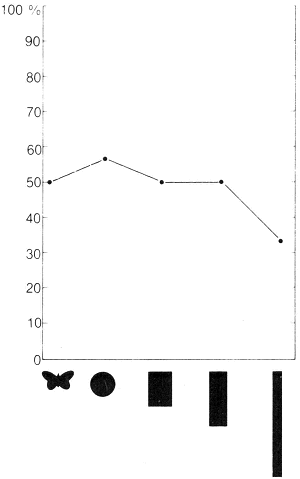
De tijdens de ‘hengelexperimenten’ gebruikte modellen. De punten in de grafiek geven het percentage positieve reacties weer, die de modellen bij mannelijke heidevlinders opriepen.
een bepaalde, constant gehouden afstand werden aangeboden en daar zonder dichterbij te komen op en neer werden bewogen. Als wij dit deden met een model van ongeveer normale grootte en een dat bijna vier maal zo groot was, kregen wij, afhankelijk van de afstand, verschillende resultaten. Op een afstand van 50 cm wekte het grote model ongeveer half zoveel reacties op als het kleine en in veel gevallen joeg het het mannetje weg. Op 100 cm afstand vertoonde het mannetje geen spoor van angst en wekte het grote model meer reacties op dan het standaardmodel!
Nu prikkelt een groter model, wanneer men het vanaf een zelfde afstand aanbiedt als een kleiner model, natuurlijk een groter deel
| |
| |
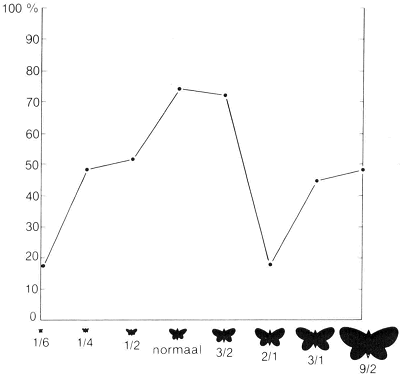
In dit experiment werden modellen van gelijke vorm, maar verschillende afmeting gebruikt. De grafiek toont het percentage positieve reacties dat ieder model opriep als men het in de richting van een mannelijke heidevlinder bewoog.
van het netvlies. Was de hogere signaalwaarde hieraan toe te schrijven of zou het verschil ook blijven bestaan als de twee modellen vanuit een gelijke hoek zouden worden gezien? Om dit na te gaan voerden wij twee series experimenten uit; één met een model van 4 cm doorsnede dat wij op 50 cm afstand aanboden en een model van 8 cm middellijn dat wij op een meter afstand hielden, en één reeks met modellen van 2 en 4 cm middellijn, die wij vanaf respectievelijk 30 en 60 cm afstand aanboden. Bij beide reeksen experimenten wekte het grotere model aanzienlijk meer reacties op dan het kleine. Dit toonde ons tegelijkertijd dat onze vlinders in staat waren om afstand te schatten, met andere woorden dat zij ‘diepte’ konden zien.
Wij zetten deze experimenten niet voort door steeds grotere modellen aan te bieden, zodat wij niet weten of wij het resultaat moeten omschrijven als ‘hoe groter, hoe beter’, maar het feit dat een dub- | |
| |
bel zo groot model beter voldeed dan een model van normale afmetingen, was op zichzelf opvallend genoeg.
Ik ga niet proberen om alle andere experimenten die wij deden te beschrijven, maar enkele van de meest belangrijke resultaten moeten toch wel even vermeld worden. Als wij een en hetzelfde model vanaf verschillende afstanden aanboden, en het op de plaats zelf op en neer lieten dansen, ontdekten wij dat het des te meer reacties uitlokte naarmate het dichter bij de mannetjes gehouden werd. In andere proeven gingen wij na in hoeverre de manier waarop het model bewoog van invloed was. Wij bewogen een ronde schijf ofwel zachtjes horizontaal ofwel met een dansende beweging zoals een vlinder in zijn vlucht op en neer danst, ofwel wij lieten het model snel rond zijn as draaien, op die manier op overdreven wijze het fladderen van een vlinder imiterend, waarbij nu eens de brede, dan weer de smalle kant van het model zichtbaar was. Zowel de dansende als de draaiende schijf wekten tweemaal zoveel reacties op als het zachtjes heen en weer bewegende model.
Wij ontwierpen andere experimenten om de interactie van de verschillende effectieve prikkels te bestuderen: de donkere tint van het model, afstand, afmeting en beweging, en de algemene conclusie die wij hieruit konden trekken, was dat een te weinig van de ene prikkel (bijvoorbeeld te lichte kleur) gecompenseerd kon worden door een extra dosis van een andere factor (bijvoorbeeld afstand). Hieruit bleek de merkwaardig automatische aard van dergelijke reacties. Men zou wellicht kunnen verwachten dat een abnormaal kleurig model des te minder reacties zou opwekken naarmate de afstand tot het mannetje kleiner werd; het tegengestelde had echter plaats: hoe ‘slechter’ een model, des te dichterbij moest het komen om reacties op te wekken.
Deze waarnemingen roepen een aantal vragen op en vormen aanleiding voor een reeks van andere experimenten. Bijvoorbeeld: zijn de zwarte modellen effectiever omdat zij feller afsteken tegen de omringende lucht? Zouden de witte modellen misschien als de beste uit de bus komen als men ze tegen een zwarte achtergrond zou aanbieden? Wat is het effect van de draaiende en dansende beweging? Is het alleen de snelheid van partiële bewegingen in beide gevallen en de afwisseling, op ieder deel van het netvlies, van licht en donker? Op deze vragen bestaat tot op vandaag nog geen antwoord; in feite beginnen wij nog maar net te ontdekken wat de algemene problemen zijn die door dergelijk onderzoek aan het licht komen. Een daarvan is bijvoorbeeld: hoe worden deze prikkels, zoals wij die tij- | |
| |
dens ons onderzoek ontdekt hebben, gebruikt? Het is duidelijk dat elk van deze zogenaamde prikkels ingewikkelde, chemische processen in gang zetten.
Een andere vraag heb ik al gesteld: hoe wordt voorkomen dat bepaalde sensorische prikkels adequate reacties opwekken? Het is heel goed mogelijk dat wij hier de uiterste grens van veldonderzoek hebben bereikt en dat deze problemen in het laboratorium moeten worden uitgezocht. Wat momenteel ook inderdaad gebeurt.
| |
Zeg het met een geurtje
De seksuele achtervolging vormt maar één schakel uit het gehele baltsgedrag van de heidevlinder. Wij begonnen het onderzoek naar de andere schakels door modellen van wijfjes aan te bieden, die vastgeprikt waren op ijzerdraadjes, nadat wij de mannetjes eerst door vliegende modellen naar de grond hadden gelokt. Helaas is dit onderzoek nooit helemaal voltooid, hoewel wij het tien seizoenen lang hebben voortgezet. Niettemin hebben wij toch een aantal interessante zaken ontdekt. Zo blijft bijvoorbeeld het mannetje tijdens zijn baltsritueel voortdurend geconfronteerd met een wijfje dat volkomen roerloos op haar plaats blijft zitten en toch vormde ieder onderdeel van zijn gedrag een reactie op verschillende, van haar uitgaande prikkels. Sommige schakels van het gedrag werden opgewekt door visuele prikkels, andere evenzeer door de geur van het wijfje. Verder waren de antennen van het wijfje onmisbaar voor de oriëntatie van het mannetje als hij zich recht voor haar posteerde. Wij konden dit aantonen door een dood wijfje aan te bieden waarbij de antennen verwijderd waren en twee spelden bij wijze van antennen in haar achterlijf geprikt waren. Het mannetje ging dan achter zo'n model staan in plaats van ervóór. Toch oriënteerde hij zich niet op de antennen wanneer hij om het wijfje heen liep om met haar te paren en hij copuleerde met een model dat ‘antennen’ aan het verkeerde uiteinde van het lichaam droeg, in de normale positie. Het meest opzienbarende onderdeel van het baltsgedrag van het mannetje vormde het ‘buigen’ en wij vroegen ons af wat de functie daarvan zou kunnen zijn. Het gedrag van het wijfje verschafte ons daarover opheldering. Wanneer het mannetje buigt, strekt het wijfje haar antennen naar hem
uit en de uiteinden ervan raken zijn voorvleugels, waarop het mannetje de antennen tussen zijn vleu- | |
| |
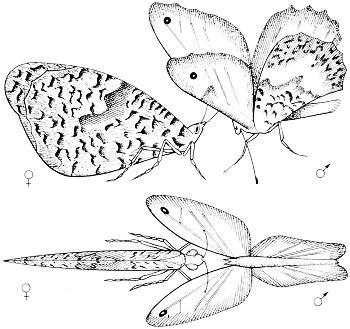
De ‘baltsdans’ van de mannetjes-heidevlinder, van opzij en van bovenaf gezien. De antennen van het wijfje betasten de geurschubben aan de binnenzijde van de vleugels van het mannetje.
gels drukt. Soms tolereerde een wijfje de capriolen van het mannetje een tijd lang, maar vloog ze weg als hij met haar wilde paren. De weigering volgde altijd precies op het moment dat de knotsvormige uiteinden van de antennen de voorvleugels van het mannetje raakten. Nu liggen juist in deze knotsvormige uiteinden de reukzintuigen. Terwijl wij de buiging van het mannetje nauwlettend gadesloegen, zagen wij dat de antennen van het wijfje de vleugels van het mannetje raakten op de plaats waar bij hem een donkergekleurde vlek voorkwam, die bij de wijfjes ontbrak. Onder de microscoop zagen wij dat dit gedeelte van de vleugel bestond uit afwijkend gevormde schubben; deze waren niet plat en breed, maar smal, en eindigden in een minuscuul kwastje. Dit type schubben is heel bekend en men treft het bij talrijke soorten Lepidoptera aan, waar het altijd gekoppeld wordt aan de produktie van geurstoffen. Dikwijls is deze geur heel duidelijk, zelfs voor de ongevoelige menselijke reuk en bij sommige soorten zelfs zeer aangenaam. De vooraanstaande bioloog Fritz Müller die als eerste deze geurschubben beschreven heeft, vermeldt hoe hij gedurende zijn reizen door Brazilië dikwijls een Papilio grayi meenam, enkel en alleen om de geur van dit dier op te snuiven als de moed hem in de schoenen zonk.
| |
| |
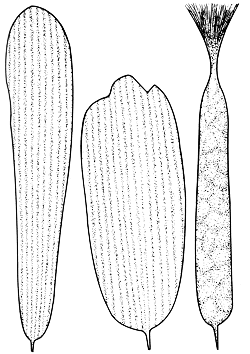
Een geurschub, voorzien van een kwastvormig uiteinde, en twee normale schubben. De foto op pag. 160 (150 maal vergroot) laat zien hoe deze schubben gerangschikt liggen op de vleugel van een mannetjesheidevlinder.
Van de heidevlinder wordt gezegd dat hij enigszins naar chocolade ruikt, maar wij durfden nooit met zekerheid te zeggen dat wij iets dergelijks roken. Maar aangezien alles er duidelijk op leek te wijzen dat de buiging van het mannetje diende om een chemische prikkel aan het wijfje aan te bieden, besloten wij dit nader te onderzoeken. Wij ontdekten al gauw dat dit gemakkelijker gezegd dan gedaan was en de kosten en moeiten die wij aan dit experiment hebben besteed, stonden vermoedelijk in geen enkele verhouding tot het magere resultaat. Wij startten ons onderzoek met het vangen van een groot aantal mannetjes en wijfjes, zo vroeg mogelijk in het seizoen. De helft van het aantal mannetjes werd onderworpen aan een onschuldige operatie: met een fijn borsteltje werden hun geurschubben verwijderd en het gedeelte van de vleugels dat zij hadden bedekt werd overgelakt met een alcoholhoudende schellak-oplossing. De overige mannetjes ondergingen een overeenkomstige behandeling, maar dan op een gedeelte van de vleugels dat juist naast het geurveld, dat
| |
| |
| |
| |

Het omlijnde gebied op de vleugel van Pararge megera geeft de ligging van de geurschubben aan. De antennen van het wijfje maken contact met deze geurvelden wanneer het mannetje zijn baltsdans uitvoert.
zelf intact bleef, gelegen was. Al deze mannetjes werden daarop vrijgelaten in de kooi en wij lieten half zoveel wijfjes bij hen toe. In de hoop dat zij onder deze omstandigheden zouden paren, keken wij toe of de wijfjes enige voorkeur zouden vertonen. Het weer dat gewoonlijk nogal wisselvallig was en de moeilijkheid om voldoende maagdelijke wijfjes te vinden, vertraagden het experiment aanzienlijk maar geleidelijk aan verzamelden wij toch gegevens van zo'n zevenentwintig paringen die wij in de kooi waarnamen. Hiervan hadden er zestien plaats met mannetjes waarvan de geurvelden intact gelaten waren en elf met mannetjes bij wie wij deze hadden verwijderd. Dit vormde een mager en niet erg significant resultaat. Wij hadden echter gezien dat verschillende van deze laatste elf paringen op een ongewone manier hadden plaatsgevonden. Een wijfje werd het hof gemaakt door een geurend mannetje dat voor haar stond, toen plotseling een niet-geurend mannetje op haar toeliep en snel met haar paarde. Dit gebeurde in de hoeken van de kooi waar de heidevlinders samengroepten. Het leek daardoor alsof de geurloze mannetjes soms door de wijfjes werden geaccepteerd als zij eerst door normale mannetjes opgewonden waren.
Onze volgende experimenten voerden wij nu in twee afzonderlijke kooien uit. In de ene kooi bevonden zich geurloze mannetjes, in de andere de geurende. Dit leverde ons een score van negen paringen tegen twee. In totaal scoorden wij dus vijfentwintig paringen tegen dertien en omdat wij wisten dat op zijn minst vijf van deze dertien paringen ‘gekaapte copulaties’ waren, staat het vrijwel vast dat de geurschubben de paring vergemakkelijken doordat zij ervoor zorgen dat het wijfje het mannetje accepteert. Het falen van de geurloze mannetjes was absoluut niet te wijten aan enige nadelige invloed
| |
| |
van de ondergane operatie op hun baltsgedrag. Als er al een verschil in gedrag bestond, was het eerder zo dat zij zich nog meer uitsloofden dan de normale mannetjes, waarschijnlijk omdat zij steeds gedwarsboomd werden door de weigering van het wijfje.
Dit resultaat, hoe bescheiden ook, verschafte ons veel genoegen, omdat hieruit een functie van de geurschubben bleek die afweek van wat men gewoonlijk aanneemt. Bij veel soorten dienen de geurstoffen om de andere sekse aan te lokken. Hier kwamen de seksen bijeen doordat het mannetje reageerde op het zien van een voorbijvliegend wijfje en begon de geur pas in de laatste fase van de balts een rol te spelen; zij diende om de medewerking van het wijfje bij de eigenlijke paring te verkrijgen.
De heidevlinder had ons dus beslist niet teleurgesteld in onze verwachtingen, maar verschafte ons een overvloed aan materiaal voor nader onderzoek. Een groot deel van ons werk werd, nadat het onderbroken was door de oorlog, niet hervat; dit was de reden dat wij ons onderzoek naar het gedrag van het mannetje niet verder voortzetten; evenmin werden microscopische onderzoekingen van de geurklieren en de olfactorische organen in de antennen, en een morfologisch onderzoek van de mannelijke en vrouwelijke copulatieorganen van de heidevlinder en verwante soorten voltooid. De onvolledigheid van de resultaten is niet de schuld van de heidevlinder, maar die van ons; en een hervatting van het onderzoek aan de heidevlinder is een van die plannen die men hoopt nog eens ooit te kunnen verwezenlijken.
| |
Aanbevolen literatuur
| Wickler, W.: Mimicry, valse berichtgeving in de natuur, Amsterdam 1968. |
| Wickler, W.: Nabootsing, camouflage en mimicry, in Grzimeks Het leven der dieren, Deel XV, blz. 138-161, Utrecht/Antwerpen 1975. |
| Frisch, O. von: Camouflage bij dieren, Den Haag 1973. |
| Burkhardt et al.: Dier en signaal; oriëntatie en communicatie in de natuur, Amsterdam 1969. |
| Fogden et al.: Dieren en hun kleuren, Amsterdam 1975. |
| Jørgensen, B.: Dieren als uitvinders, Den Haag 1969. |
| Tinbergen, N.: Het dier in zijn wereld, Deel I, Utrecht/Antwerpen 1976. |
|
|