|
| |
| |
| |
2. Meer over de bijenwolf
Ik was gedwongen om mijn onderzoek van de graafwespen twee zomers, die mijn vrouw en ik doorbrachten aan de oostkust van Groenland, te onderbreken. Tijdens de terugreis uit het noordpoolgebied, gingen mijn gedachten vaak terug naar Philanthus. Ik had het gevoel dat mijn tot dusverre verkregen resultaten slechts een pover begin vormden en ik bleef naar mogelijkheden zoeken om dieper in het vraagstuk van hun oriëntatievermogen door te dringen. De kans om op grotere schaal veldwerk te verrichten deed zich voor, toen mijn professor, H. Boschma, die inzag dat er tijdens veertien officiële vakantiedagen die een assistent waren toebedeeld niet veel werk verricht kon worden, mij toestond enige studenten mee te nemen en het werk dat zij voor mij zouden doen, als een onderdeel van hun opleiding te beschouwen.
Mijn enthousiaste verhalen over de wespen en hun prestaties hadden de verwondering van sommige van mijn vrienden op het laboratorium gewekt, zodat zij maar al te graag met mij mee wilden gaan. De waarnemingen die hieronder worden beschreven, zijn dan ook met hun enthousiaste hulp gedaan. Ons team bestond, afgezien van mijzelf, uit W. Kruyt, D.J. Kuenen, R.J. van der Linden en G. van Beusekom.
| |
De ‘oriëntatievlucht’
Wij begonnen met een nadere bestudering van de oriëntatievlucht van de wespen. Zoals ik al eerder vermeldde, beschikten wij reeds over zeer positieve aanwijzingen dat er werkelijk sprake was van echte oriëntatievluchten, maar het onomstotelijke bewijs hiervoor was nog altijd niet geleverd. De overigens irriterende grillen van het wisselvallige Atlantische klimaat boden ons een unieke gelegenheid om dit bewijs te leveren. Lange perioden van koud, regenachtig weer zijn niet ongebruikelijk voor een Hollandse zomer; in feite komt dit weertype zelfs meer voor dan de zonnige dagen die de wespen tot activiteit konden verlokken. Het regenachtige weer stelde het moreel van ons kamp nogal op de proef, maar de eerste tekenen van weersverbetering ontketenden gewoonlijk een koortsachti- | |
| |

‘Wandelende’ zandduintjes, opgeworpen door de overheersende winden te Hulshorst.
ge activiteit, waarbij ieder van ons zijn uiterste best deed, klaar te zijn voordat de wespen hun vluchten zouden hervatten.
Wij hadden al eerder opgemerkt dat vele (hoewel niet alle) wespen de koude en natte perioden in hun holletjes doorbrachten. Regen en wind richtten dikwijls schade aan onder hun bakens en wellicht vergaten de wespen zelf ook de juiste positie ervan tijdens hun ondergrondse verblijf. Hoe dan ook, zodra het weer zich herstelde, maakten alle wespen extra langdurige oriëntatievluchten als ze zich voorbereidden op hun eerste tocht. Zouden ze zich misschien de ligging van hun gebied opnieuw moeten inprenten?
Op een van die ochtenden, terwijl de grond nog nat was van de dauw maar het weer er zonnig en veelbelovend uitzag, waren wij al
| |
| |
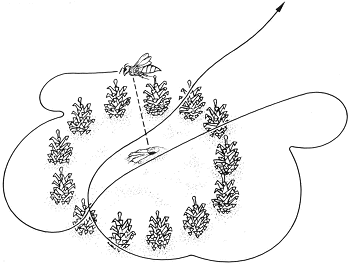
De vliegroute van een Philanthus-wesp bij het verlaten van haar nest: een typische oriëntatievlucht.
om half acht present in de kolonie. Ieder van ons installeerde zich in de buurt van een aantal nesten en lette nauwkeurig op de eerste tekenen van te voorschijn kruipende wespen. Wij hoefden niet lang te wachten voor wij bij een van de ingangen het zand, waarmee deze was afgedekt, zagen bewegen: het teken dat een wesp probeerde zich naar buiten te werken. Snel legden wij een cirkel van denneappels rondom de nestingang. Toen de wesp uiteindelijk uit het nest kwam, begon ze direct te graven en haar holletje in orde te brengen, waarna zij zand over de ingang harkte en wegvloog. In de loop van de ochtend kwam een groot aantal wespen naar buiten en elk van hen kreeg een cirkel van denneappels rond haar nestingang voordat zij daadwerkelijk ‘de deur had geopend’. Enkele van deze wespen lieten het nest voor wat het was en vlogen direct weg als ze buiten kwamen. Deze laatste wespen besloten wij te gaan gebruiken voor onze experimenten. Zoals verwacht, maakten zij uitgebreide oriëntatievluchten, waarbij ze steeds wijdere cirkels beschreven, steeds hoger boven de grond vliegend, alvorens definitief te vertrekken. Wij noteerden zorgvuldig de duur van deze vluchten. Zodra een van de wespen echt vertrokken was, namen wij de denneappels weg. Dit deden wij om er absoluut zeker van te zijn dat, wanneer een wesp onverwacht terug zou keren, zij geen denneappels rondom haar nest zou zien. Wanneer wij een wesp zagen terugkeren
| |
| |
met een bij, deden wij een verplaatsingstest door de cirkel van denneappels op enige afstand van het nest te leggen. Als de wesp zich door deze kegels liet leiden, zouden wij hebben bewezen dat zij zich de bakens moest hebben ingeprent tijdens haar oriëntatievlucht, want zij zou ze op geen enkel ander tijdstip hebben kunnen zien.
Niet alle wespen keerden diezelfde dag terug. Hun langdurig verblijf ondergronds en hun lange periode van vasten noodzaakte hen waarschijnlijk om zich eerst op de heide te voeden. Enkele keerden echter terug met een bij, en met deze exemplaren konden wij enkele schitterende experimenten doen. In totaal testten wij dertien wespen. Wij lieten hen 93 maal kiezen tussen hun echte nest en een ‘fopnest’, omgeven door een ring van denneappels. Zeventigmaal kozen zij de fopnesten, slechts twintigmaal hun echte nestingang. Bij de controle-experimenten, die wij na de echte tests namen, viel slechts drie van de 39 maal de keuze op de fopnesten, in alle andere gevallen kozen de wespen weer voor het echte nest, waaromheen wij de denneappels hadden teruggelegd. Het was nu absoluut zeker dat de wespen de aard en de positie van de nieuwe bakens tijdens hun oriëntatievlucht hadden leren kennen.
De meest indrukwekkende prestatie werd geleverd door wesp nr. 179. Ze had een oriëntatievlucht gemaakt van krap zes seconden en was daarop vertrokken zonder nogmaals terug te komen, laat staan te landen. Toen ze meer dan een uur later bij haar terugkomst werd getest, koos ze twaalfmaal de ring denneappels en kwam ze nooit in de buurt van haar echte nest! Nadat wij de oorspronkelijke situatie weer hadden hersteld, landde ze direct bij haar nest en glipte naar binnen. Deze recordprestatie werd bijna geëvenaard door nr. 174 en 177; beide wespen bleken na een ononderbroken oriëntatievlucht van dertien seconden perfect getraind. Alle andere wespen maakten ofwel langere oriëntatievluchten of onderbraken deze door een of meer keren weer op het nest te landen alvorens opnieuw te vertrekken. Dergelijke wespen leerden misschien eerder tijdens hun landing dan tijdens de rondvluchten zelf, zodat hun resultaten minder overtuigend waren.
Dit resultaat, hoewel niet geheel onverwacht, maakte desondanks toch diepe indruk op ons. Het maakte in ieder geval duidelijk dat deze kleine insekten een verbazingwekkend leervermogen bezaten en nog meer verbaasde het ons dat een wesp, als zij zich niet volledig met haar omgeving vertrouwd achtte, steeds een dergelijke oriëntatievlucht maakte, alsof het dier er zich van bewust was, wat het effect van dit speciale gedrag zou zijn.
| |
| |
Waarschijnlijk vertoonden de wespen deze oriëntatievluchten omdat zij door het lange verblijf ondergronds het resultaat van hun laatste vlucht waren vergeten. Maar de wespen wisten ook het hoofd te bieden aan de toevallige wijzigingen in de situatie op korte termijn, veroorzaakt door een schaap of misschien een ree, die de bakens uit hun positie hadden gebracht. Wij onderzochten het effect van twee vormen van verstoring op de oriëntatievluchten. In de experimenten van het type A voegden wij een niet al te opvallend baken toe aan de reeds aanwezige of haalden er juist een weg, voordat de wesp terugkeerde en herstelden dan de oorspronkelijke situatie weer als zij in haar nest was. Deze wespen maakten, hoewel zij de oude bekende situatie aantroffen bij het verlaten van hun nest, lange oriëntatievluchten. Bij de experimenten van het type B werden de wespen totaal ongemoeid gelaten als zij hun nest binnengingen, maar werden dezelfde veranderingen aangebracht als in de experimenten A als zij het nest weer verlieten. Deze wespen maakten geen van alle oriëntatievluchten. De wespen in de experimenten van type A vertoonden steeds enige aarzeling alvorens te landen. Met andere woorden: veranderingen in de vertrouwde omgeving aangebracht op het ogenblik van terugkeer, stimuleerden de wesp tot het maken van een oriëntatievlucht bij haar eerstvolgende vertrek, terwijl diezelfde verstoringen, aanwezig op het tijdstip van vertrek, geen effect hadden!
Enkele andere, tamelijk onvolledige experimenten vestigden onze aandacht op een ander interessant facet van de zaak. Opvallende nieuwe bakens, aangebracht vóór de terugkeer van de wesp en na haar vertrek daar gelaten, beïnvloedden zowel de aard als de duur van de oriëntatievlucht: de wesp cirkelde in zo'n geval altijd een paar keer rond dit speciale baken. Wanneer men een dergelijk baken enige tijd op zijn plaats liet liggen, zodat de wesp het verschillende malen tijdens haar aankomst en vertrek kon zien, en het vervolgens verplaatste, maakte de wesp gewoonlijk een wat langere oriëntatievlucht, maar zonder extra rondjes boven het speciale baken te vliegen. Ze herkende het baken duidelijk en hoefde zich alleen maar de nieuwe positie ervan in te prenten. Deze experimenten waren te gering in aantal en niet honderd procent overtuigend, maar zij lieten wel vermoeden dat het hele verschijnsel oriëntatievlucht meer omvatte dan wij aanvankelijk gedacht hadden. Het hele merkwaardige fenomeen verdiende ongetwijfeld verder onderzoek.
| |
| |
| |
De aard van de bakens
Vervolgens gingen wij ons afvragen wat de wezenlijke aard was van de bakens die de wespen gebruikten. Wat prentten zij zich nu precies in? Wij besteedden verscheidene seizoenen aan de beantwoording van deze vraag en de meest opmerkelijke van de uitgevoerde experimenten zijn waard om hier beschreven te worden.
Allereerst ontdekten wij dat niet alle voorwerpen van even grote betekenis waren voor de wespen. De eerste aanwijzing hiervoor kregen wij toen wij probeerden, hen te trainen op velletjes gekleurd papier van 7 × 10 centimeter, die wij rondom de nestingang neerlegden, als een soort inleiding op een onderzoek naar het vermogen om kleuren te zien. Het bleek vrijwel onmogelijk om de wespen ertoe te bewegen zelfs een combinatie van drie van dergelijke velletjes papier als baken te gebruiken; zelfs nadat wij ze dagenlang onafgebroken op dezelfde plaats hadden laten liggen, boekten wij nauwelijks resultaat met dezelfde eenvoudige verplaatsingstest die zo uitstekend voldeed bij de denneappels. De meeste wespen keken niet eens naar de papiertjes. Toch hadden wij de indruk dat de helder blauwe, gele en rode velletjes zeer opvallend moesten zijn. Om de een of andere reden voldeden de denneappels beter aan de eisen die de wespen aan bakens stelden dan de vlakke velletjes papier.
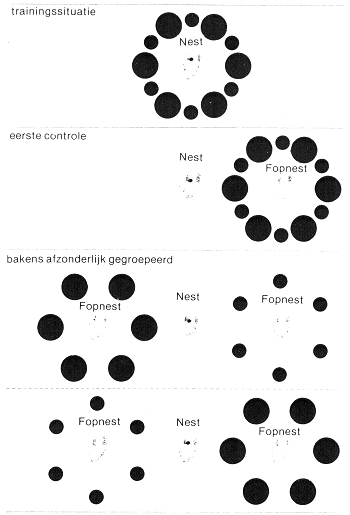
Samen met Kruyt bedacht ik een experiment om dit te toetsen. Wij legden twee typen voorwerpen rondom een nest, bijvoorbeeld platte schijven en denneappels, afwisselend in een cirkel gerangschikt. Na een dag of wat verplaatsten wij de hele cirkel en keken of de wesp zich erdoor liet leiden. Als dat het geval was, maakten wij twee fopnesten, een aan iedere zijde van het echte nest, even ver er vandaan, en legden alle voorwerpen van hetzelfde type rondom een van deze fopnesten, alle voorwerpen van het andere type rond het andere fopnest. Wanneer de wesp zich vooral op een van beide typen objecten had getraind, zou zij een van beide fopnesten de voorkeur geven. Een dergelijk gedrag zou nergens anders aan kunnen worden toegeschreven dan aan een verschil in instelling van de wesp ten opzichte van de twee typen bakens, want beide konden even vaak door de wesp zijn opgemerkt, aangezien hun afstand tot de nestingang gelijk geweest was en zij allemaal rondom deze ingang waren aangeboden; kortom, beide typen voorwerpen hadden evenveel kans om gekozen te worden.
Op deze wijze vergeleken wij vlakke voorwerpen met driedimensionale, donkere met lichte bakens die contrasteerden met de achter- | |
| |

Dit experiment dient om de voorkeur voor bepaalde bakens te testen. Een wesp wordt geconfronteerd met zowel grote, platte schijven als kleine, massieve bolletjes, die beide als baken rondom het nest worden geplaatst. Vervolgens groepeert men de twee typen bakens ieder afzonderlijk rond fopnesten aan weerszijden van het werkelijke nest (zie tekening blz. 41). Nu streek de wesp steeds in het centrum van de cirkel ronde bollen neer, wat aantoont dat deze de belangrijkste bakens vormen.
grond met bakens die er juist mee harmonieerden, grotere bakens met kleinere, dichtbijgelegen objecten met verder verwijderde, etc. Iedere test diende gedaan te worden met talrijke wespen en iedere wesp moest meermalen kiezen voor wij er zeker van konden zijn dat haar voorkeur echt constant bleef. Dit vergde zeer veel tijd, maar de resultaten waren de moeite waard. De voorkeur die de wespen voor bakens vertoonden, verschilde werkelijk van de onze. Als wij platte ronde schijven en halve bollen met dezelfde diameter aanboden, kozen de wespen altijd de halve bollen (43 scores voor, 2 tegen). Dit was niet een gevolg van het grotere oppervlak van de halve bollen, want als wij de proef herhaalden met platte schijven met een veel grotere diameter (tien centimeter middellijn, terwijl de doorsnede van de halve bollen maar vier centimeter was) waren de scores nog 73 voor, 19 tegen, ten gunste van de halve bollen.
| |
| |
Tijdens andere experimenten ontdekten wij dat de halve bollen niet de voorkeur genoten vanwege hun schaduwwerking, noch omdat zij contrast vertoonden tussen helderverlichte en diepzwarte vlakken, noch omdat zij driedimensionaal waren, maar vanwege het feit dat zij boven de grond uitstaken. Het doorslaggevende bewijs hier- | |
| |
voor werd geleverd door holle kegels aan te bieden, die voor de helft gewoon op de grond gezet waren en voor de andere vijftig procent ondersteboven in de bodem waren ingegraven. Beide typen bakens waren driedimensionaal, maar het ene type stak boven de grond uit, terwijl het andere putjes in de bodem vormde. Vrijwel zonder uitzondering kozen de wespen voor de rechtopstaande kegels (108 tegen 21 scores).
De voorkeur voor voorwerpen die boven de grond uitstaken, was een van de redenen waarom denneappels zo gewild waren als baken. Een andere reden was dat de dennekegels een regelmatig patroon van lichte en donkere vlekjes vertoonden, terwijl zij in de derde plaats een ruw in plaats van een glad oppervlak hadden, d.w.z. dat er van ingesneden voorwerpen een grotere aantrekkingskracht uitging dan van gladde. Andere studenten waren tot soortgelijke bevindingen gekomen in een onderzoek aan honingbijen en dit houdt waarschijnlijk voor een groot deel verband met de structuur van de samengestelde ogen van insekten.
Verder ontdekten wij dat grote voorwerpen geschikter waren dan kleine, dichtbijgelegen objecten beter dan diezelfde voorwerpen, maar dan verder weg gelegen, voorwerpen die contrasteerden met de achtergrond beter dan objecten die ermee harmonieerden, voorwerpen gepresenteerd tijdens kritieke fasen (zoals bij het begin van het graven van een nieuw nest of onmiddellijk na een periode van regen) geschikter dan voorwerpen die men aanbood als een wesp eenmaal een bepaalde kennis van zijn omgeving had.
Terwijl wij deze experimenten deden, verwonderde het ons dikwijls dat de wespen vaak zo gauw een fopnest kozen, hoewel de aangeboden cirkel slechts half zoveel voorwerpen telde als die waarop de dieren waren getraind. Dit zou niet zo vreemd geweest zijn als de wespen deze ‘zwakkere’ bakens eenvoudig hadden genegeerd, maar dit was niet het geval. Wanneer wij bij onze oorspronkelijke proeven met platte schijven en halve bollen alleen de schijven aanboden, kozen de wespen, geconfronteerd met de keuze tussen de schijven en hun eigen nest zonder schijven of halve bollen, vaak de schijven. Deze werden dus niet volledig genegeerd; zij vormden potentiële bakens, maar waren als zodanig minder van betekenis dan de halve bollen. Toen wij dit eenmaal wisten, ontdekten wij dat wij met enige volharding de wespen ook konden trainen op onze stukjes papier. Maar het duurde lang.
| |
| |

Een ander experiment omtrent het gebruik van bakens: men bood de wespen zowel lange staande blokjes, als korte maar grotere blokjes aan. De wesp koos gewoonlijk de langste bakens uit.
| |
De eerste vorderingen
Op grond van het feit dat de wespen deze cirkels, die slechts uit half zoveel elementen bestonden als zij gewoon waren te zien, accepteerden, vermoedden wij dat zij zowel op de cirkelvormige rangschikking zelf als op de eigenschappen van de afzonderlijke voorwerpen letten. Dit zou betekenen dat hier sprake was van ‘vorm-afhankelijke’ stimuli en het leek goede experimentele mogelijkheden te bieden. Het was Van Beusekom die zich hiermee ging bezighouden en die door middel van een aantal vernuftige experimenten aantoonde dat de wespen inderdaad reageerden op een zeer gecompliceerde prikkelsituatie.
Om te beginnen stelde hij vast dat de wespen bakens zoals denneappels tamelijk goed konden herkennen. Hij trainde de wespen met de gebruikelijke cirkel van denneappels en liet ze vervolgens kiezen tussen deze cirkels en een overeenkomstige rangschikking van gladde blokjes, die ongeveer het formaat van denneappels hadden. De wespen kozen hoofdzakelijk de denneappels, waaruit bleek dat zij reageerden op kleine verschillen tussen de twee typen bakens.
Vervolgens trainde hij een aantal wespen met een cirkel van zestien denneappels en deed toen twee soorten proeven met deze dieren. In de experimenten van type A moest de wesp kiezen tussen twee arrangementen van zestien dennekegels, waarvan het ene stel een cirkel vormde, het andere een daarvan afwijkende vorm, zoals een vierkant, een driehoek of een ellips. Hij ontdekte dat de wespen in
| |
| |
staat waren onderscheid te maken tussen beide figuren, tenzij de ene figuur zeer veel leek op een cirkel, en dat zij altijd neerstreken in de cirkel. Bij deze experimenten bleken de afzonderlijke kegels niet van belang te zijn; hij kon dezelfde exemplaren gebruiken voor zowel een cirkel, een vierkant of een driehoekige figuur. De wesp koos duidelijk de hele cirkel, niet de denneappels van de training.
In de experimenten van type B bood hij, na de gebruikelijke training met een cirkel van zestien kegels, deze zestien dennekegels aan in een niet-cirkelvormige rangschikking in combinatie met acht of zelfs minder denneappels die wel tot een - vrij open - cirkel waren gerangschikt en hij zag dat de wespen toch deze laatste cirkels kozen ondanks het geringe aantal denneappels waaruit zij bestonden. Hij kon zelfs nog verder gaan en een cirkel aanbieden die uit totaal verschillende elementen bestond, zoals vierkante blokjes (die, zoals uit andere experimenten was gebleken, door de wespen goed onderscheiden werden van denneappels). Gaf hij de wespen de keuze tussen dergelijke cirkels en denneappels, die niet cirkelvormig waren gerangschikt, dan kreeg altijd de cirkel de voorkeur. Zodoende kwam het langs verschillende wegen tenslotte vast te staan dat de wespen niet alleen reageerden op afzonderlijke bakens (zoals de voorkeursexperimenten van Kruyt en mijzelf hadden aangetoond), maar ook op de gehele cirkel als gegeven figuur.
| |
Verder weg gelegen bakens
Al deze experimenten hadden, hoewel ze ons waardevolle informatie gaven over de manier waarop de wespen zich een indruk van hun omgeving vormden, één beperking gemeen: ze maakten alleen duidelijk hoe de wespen zich gedroegen tijdens de laatste etappe van hun thuisreis. Wij beschikten over talrijke aanwijzingen, dat de denneappels pas werden opgemerkt als de wespen nog slechts een paar meter van hun nest waren verwijderd. Hoe waren zij er echter

| |
| |
in geslaagd om tot zover de weg te vinden?
Hoewel wij ons bewust waren van deze beperkingen, was het zeer moeilijk om onze experimenten over grotere oppervlakten uit te breiden. Toch probeerden wij het op verschillende manieren. Een paar maal verplaatsten wij kleine denneboompjes die op enige afstand van de door ons geobserveerde nesten groeiden. In veel gevallen werden de wespen hierdoor om de tuin geleid en probeerden zij hun nesten te vinden in een positie die correct was ten opzichte van de verplaatste boompjes. De precisie van hun oriëntatievermogen met behulp van dergelijke ver verwijderde bakens was werkelijk verbazingwekkend.
Van dergelijke grote bakens werd op enigszins andere wijze gebruik gemaakt dan van de denneappels. In de eerste plaats fungeerden zij ook wanneer de wesp op grotere afstand van het nest was, als baken. Ten tweede konden wij ze over veel grotere afstanden verplaatsen dan de dennekegels. Een cirkel van denneappels had geen enkel effect meer als richtingaanwijzer voor de wespen als wij deze meer dan een meter of twee verplaatsten, maar een denneboompje of zelfs een tak van ongeveer een meter hoog, kon, zelfs wanneer ze meer dan acht meter werden verzet, nog wespen op een dwaalspoor brengen. Bij veel van onze voorgaande experimenten hadden wij bovendien waargenomen dat de wespen, als ze de directe omgeving van het nest niet in de vertrouwde staat aantroffen, rondom een denneboompje of een grote zandheuvel, die wel zeventig meter verderop kon liggen, gingen cirkelen om daarna opnieuw naar het nest te vliegen. Het leek erop dat zij aan de hand van deze grotere bakens opnieuw hun richting bepaalden.
Van der Linde en anderen besteedden veel tijd en energie om afzonderlijke wespen over afstanden van maximaal een kilometer naar alle richtingen te transporteren in een doek waar geen licht doorheen drong. Aangezien de beste jachtterreinen zich ten zuiden en zuidoosten van de kolonie uitstrekten, terwijl de Philanthus-vlakte naar alle overige richtingen begrensd werd door kale zandvlakten of
| |
| |

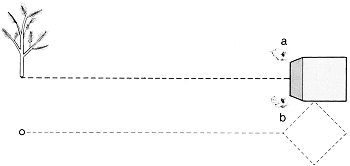
Dit experiment diende om na te gaan hoe de wespen zouden reageren op een combinatie van meerdere bakens. De resultaten worden hieronder besproken.
dichte dennenbossen, mochten wij veronderstellen dat onze wespen het gebied ten zuiden en zuidoosten van hun woongebied beter kenden dan dat in de andere windrichtingen; een vermoeden dat versterkt werd door het feit dat de wespen altijd naar het zuiden of zuidoosten wegvlogen en vandaar met een bij terugkeerden. Toen wij dan ook gingen kijken welke van de getransporteerde wespen het snelst bij hun nesten terugkeerden, bleek dat zij vanuit het zuiden en zuidoosten veel gemakkelijker de weg terugvonden dan vanuit de overige richtingen. De wespen die naar het noordwesten waren gebracht bijvoorbeeld, keerden gedurende de hele observatieperiode niet terug. Dit leek erop te wijzen dat de terugkeer vanuit onbekend gebied moeilijk, zo niet onmogelijk was, en dat dus een of andere vorm van ervaring opdoen hiervoor onontbeerlijk was; meet kon het experiment ons daarover echter niet zeggen.
Een van de testen gaf ons een goede indruk van de manier waarop de wespen gebruik maakten van combinaties van bakens. Een be paalde wesp kreeg een vierkant blok van zwart hout aangeboden
| |
| |
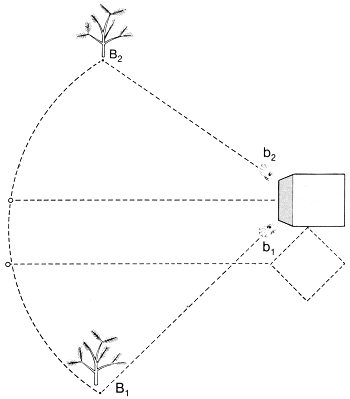
25 × 25 × 3 cm groot, dat vlak bij het nest werd neergelegd en een dennetak van iets meer dan een meter hoog, die ongeveer een meter van het nest vandaan gezet werd. Het blok raakte met een van zijn hoeken de nestingang. Toen de wesp gewend was geraakt aan deze rangschikking, verzette Van Beusekom zowel de tak als het blok ongeveer 25 cm; de wesp koos het blok en probeerde haar nest te openen bij punt X (zie tekening). Draaiden wij nu het blok 45 o, dan wist de wesp niet bij welke hoek ze moest gaan graven; ze probeerde het vijfmaal bij hoek a, vier maal bij hoek b en vijf maal ergens daar tussenin.
Verplaatsten wij daarop de dennetak zodanig dat deze in positie B1 terechtkwam, dan koos de wesp zevenmaal hoek b2 en driemaal hoek b1; zetten wij daarna de tak in positie B2, dan vloog de wesp achtmaal naar b1 en slechts tweemaal naar b2. Deze getallen zijn weliswaar klein, maar in verschillende andere experimenten konden wij dezelfde tendens waarnemen: de wespen maakten gebruik van meerdere bakens: sommige na elkaar, andere tegelijkertijd.
| |
| |
| |
De jacht
Terwijl wij zo stap voor stap steeds meer te weten kwamen over de oriëntatievermogens van Philanthus, werden wij voortdurend nieuwsgieriger naar een ander aspect van hun levenswijze: hun bijna griezelige vermogen om feilloos honingbijen te herkennen tussen de talrijke insekten op de heide, waarvan er verschillende zeer veel gelijkenis met de honingbij vertonen, zoals sommige solitair levende bijen en vooral de zweefvlieg Eristalis tenax, een soort die met zoveel succes honingbijen imiteert dat zelfs vogels zich dikwijls laten misleiden, zoals de experimenten van Mostler hebben aangetoond. Hoewel Philanthus uitstekend in de verte kan zien, zoals onze dagelijkse experimenten op het gebied van hun oriëntatievermogen aantoonden, zien zij niet erg scherp en het was dan ook absoluut uitgesloten dat zij de fijnere details in de bouw van een bij konden zien. Wij veronderstelden daarom dat een wesp tijdens de jacht van andere zintuigen gebruik zou maken. Misschien herkende zij de karakteristieke toonhoogte van het gezoem, zoals wij, en ook sommige op hommels jagende vogels, dat doen. Ook zouden reukof tastzin hierbij een rol kunnen spelen. Een andere mogelijkheid was dat Philanthus de bijen als zodanig niet eens herkende, maar op de korven reageerde en de bijen ving als deze hun korven verlieten of binnen gingen. In dit laatste geval zouden ze niet eens andere insekten tegenkomen en zou er zich helemaal geen herkenningsprobleem voordoen.
Terwijl Kruyt en Van Beusekom hun onderzoek naar de oriëntatie voortzetten op de zandvlakte, besloot ik een seizoen te gaan kijken hoe de wespen zich tijdens de jacht gedroegen. Dit was gemakkelijker gezegd dan gedaan. Er waren, en dit is niets teveel gezegd, enkele duizenden wespen. Maar een eerste vluchtig onderzoek wees uit dat zij in een uitgestrekt gebied van ongeveer anderhalve vierkante kilometer op jacht gingen. Ondanks urenlang observeren in de buurt van de dichtstbijzijnde bijenkorven, zag ik nooit een wesp in de buurt van deze korven komen en het was dus duidelijk dat ze niet bij de ingang daarvan op de bijen loerden. Ik besloot daarom mijn probleem anders aan te pakken. Tijdens onze oriënteringsexperimenten hadden wij gezien dat een wesp in staat was haar bij terug te vinden als ze hem verloren had. Sommige wespen lieten wel hun bij vallen als wij onze experimenten te lang voortzetten, maar als wij daarna de proef beëindigden en de denneappels weer vlak bij het nest teruglegden, zodat de wesp haar eigen ingang weer kon
| |
| |
vinden, ging het dier gewoonlijk eerst het nest open maken en inspecteren, om daarna haar bij op te halen. Ook liet een wesp wel plotseling haar prooi vallen als ze werd opgeschrikt door een onverwachte beweging van een van ons. Dikwijls slaagden zulke dieren er dan weer in hun bij terug te vinden, een opmerkelijke prestatie, aangezien de betreffende prooi soms metersver van het nest terechtgekomen was, tussen het donkerbruine mos waar hij zelfs voor ons vrijwel onzichtbaar was.
De manier waarop een wesp op een zoekgeraakte bij afging, leverde mij zeer veel informatie op. Ze vloog op een ongebruikelijke, opvallende wijze rond, daarbij laag boven de grond wijde cirkels beschrijvend. Plotseling hield ze haar vlucht in, zette dan, hangend in de lucht, op een eigenaardige zigzaggende manier haar vlucht weer voort, waarbij ze steeds lager boven de grond kwam en landde tenslotte op enkele centimeters van haar prooi. Daarna liep ze op haar bij toe terwijl ze haar antennen op en neer bewoog, en greep dan in minder dan een seconde de bij beet.
Het viel ons op dat de wespen bij dergelijke gelegenheden steeds hun verloren geraakte bijen tegen de wind in naderden. Het leek alsof zij tijdens hun zoekactie plotseling gealarmeerd werden door de geur van de bij die door de wind werd aangevoerd. Twee eenvoudige experimenten toonden ons dat dit inderdaad het geval was.
Wanneer ik een wesp liet schrikken zodat ze haar bij liet vallen en ik dan snel deze bij in een open buisje stopte dat ik met gaas afdekte en in de bodem ingroef, zodat de bij zelf onzichtbaar was maar haar geur wel kon ontwijken, naderde de wesp deze buisjes altijd tegen de wind in, precies zoals wanneer ze de bij helemaal kon zien. Uiteindelijk landde zo'n wesp dan op het gaas, waarover ze dan rondliep en als waanzinnig probeerde om in de buis te komen.
Een andere proef, oorspronkelijk opgezet met een heel ander doel, wees in dezelfde richting. Aangezien wij op goede gronden vermoedden dat de reukorganen van de wesp gelokaliseerd waren op de antennen, wat later ook bevestigd werd, hadden wij, pogend om duidelijk aan te tonen dat zij zich oriënteerden met behulp van hun ogen en niet via de reukorganen, bij sommige wespen de antennen verwijderd, en hen vervolgens gebruikt voor onze experimenten met de bakens. Dergelijke wespen bleven gewoon werken aan hun nesten; zij vlogen uit en maakten oriëntatievluchten, ze volgden precies onze bakens als wij die verplaatsten, maar ze brachten nooit een prooi mee naar huis. Dit sterkte ons in ons vermoeden dat de reuk een grote rol speelde bij de jacht.
| |
| |
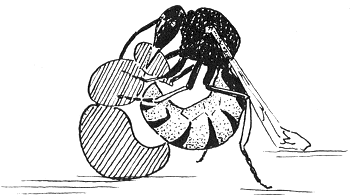
Mijn volgende poging om iets van het jagen te zien, ondernam ik eveneens vanuit de veronderstelling dat het onbegonnen werk was om zomaar het veld in te gaan en daar naar jagende wespen te zoeken. In Fabre's artikel over Philanthus had ik gelezen dat men deze wespen in gevangenschap kon leren bijen te vangen, door eenvoudig een wesp met een paar levende bijen in een jampot te zetten. Ik besloot dit experiment te herhalen. Aanvankelijk probeerde de wesp te ontsnappen, daarna kalmeerde ze, maakte haar antennen schoon, wreef haar pootjes en vleugels, maar vertoonde niet de minste aandacht voor de bijen. Nu en dan, wanneer een van de bijen tegen haar aanvloog, trok zij zich terug of draaide ze rond, alsof ze zich gereed hield voor de verdediging. Ik begon er al aan te twijfelen of ik nog iets te zien zou krijgen dat de moeite waard was. Toen greep de wesp onverwacht, vlak voor een bij haar weer zou raken, de aanvaller vast, draaide zich met de bij in haar poten om en voordat ik kon volgen wat er precies gebeurde, had zij haar prooi al onder ‘de kin’ gestoken. De bij viel onmiddellijk op haar rug, trok nog enkele seconden met haar pootjes en bleef toen bewegingloos liggen. Philanthus gaf haar slachtoffer nu een zeer eigenaardige behandeling. Zij drukte de bij stevig tegen haar lichaam, zodat er nectar uit de mondopening van het dier vloeide. Deze likte de wesp tot de laatste druppel op. Vervolgens bracht ze de bij in transportpositie (omgekeerd onder haar lichaam, vastgehouden bij het middelste paar poten) en vloog ermee op, om echter tegen het glas te botsen en weer op de tafel te belanden.
Ik herhaalde dit experiment een paar keer en kreeg zo uiteindelijk een volledig beeld van wat er gebeurde. De wesp greep een bij alleen, wanneer deze haar antennen raakte. Ze draaide het dier dan
| |
| |
bliksemsnel om, totdat het met zijn kop naar haar toegewend was en terwijl de ongelukkige bij als een razende probeerde zijn belaagster te steken, waarin zij zelden slaagde omdat de angel afketste op het harde pantser van Philanthus, wist de wesp met feilloze precisie de kin van de bij te vinden en daar haar vervaarlijke angel in te steken.
Dit drama-in-miniatuur leverde mij een aanzienlijke hoeveelheid informatie. Het feit dat de aanraking van de antennen het volledige jachtgedrag ontketende, toonde duidelijk aan dat het een of andere op de antennen gelegen zintuig hierbij een rol speelde. Iedereen die wel eens antennen van een insekt onder een microscoop heeft bekeken, weet dat deze bedekt zijn met vele duizenden kleine structuurtjes van allerlei aard, alle verbonden met zenuwuiteinden, die op hun beurt weer in verbinding staan met de grote antennezenuw. Deze structuren, allemaal zintuigen, vervullen onmiskenbaar een groot aantal functies. Er zijn er minstens een paar bij met een tastof reukfunctie. Ik deed een eenvoudige proef om uit te maken welk van deze zintuigen in dit speciale geval een rol speelde. Ik stopte tegelijk met honingbijen enkele bromvliegen en hommels in de jampot. Zelfs als deze insekten in contact kwamen met de antennen van de wesp, werden ze ongemoeid gelaten. Toch bewees dit nog niet alles: zij zouden op grond van tastzinutuigen kunnen worden onderscheiden. Nu nam ik een paar kleine hommels en schudde deze in een glazen buis samen met levende bijen, om ze de geur van bijen te laten aannemen. Tot mijn tevredenheid zag ik dat mijn wespen nu wel belangstelling begonnen te tonen voor deze ‘geparfumeerde’ hommels, een soort chemische imitatiebijen eigenlijk, en dat zij zelfs een ervan vingen en staken. Dit toonde op zijn minst aan dat chemische stimuli een rol speelden, althans zeker onder deze tamelijk onnatuurlijke omstandigheden waaronder ik deze experimenten deed.
Ik besefte echter dat deze proeven mij nog niet zoveel verder hadden gebracht. De wespen die zoekgeraakte bijen opspoorden, zouden heel goed op een andere manier te werk kunnen gaan dan wanneer ze op jacht waren (en inderdaad ontdekte ik later dat dit zo was). De wespen in de jampot waren trouwens helemaal niet op jacht; ze wilden alleen maar reageren als de bijen zich, bij wijze van spreken, aan hen opdrongen.
| |
| |
| |
Naar de echte jachtgebieden
Er bleef mij niets anders over dan toch maar naar het echte jachtterrein te trekken. Het zomaar ergens gaan zitten in de uitgestrekte heidevlakte en dan maar te wachten op bijenvangende wespen in mijn buurt, leek een tamelijk hopeloze onderneming. Zou ik geen stukje terrein kunnen vinden waar de wespen zich waarschijnlijk zouden concentreren? Ik beredeneerde dat de zoom van de heide, die het dichtst aan de kolonie grensde, de meest geschikte plaats moest zijn, vooral wanneer de wind uit zuidoostelijke richting woei en de wespen niet zo ver zouden willen vliegen. Dus installeerde ik mij een paar dagen aan de noordwestelijke rand van de heide en was inderdaad zo gelukkig om een paar maal het complete jachtgedrag te kunnen observeren. Verschillende malen per dag zag ik felgele wespen over de heide vliegen. Sommige peurden honing uit de Calluna-bloempjes, andere streken van tijd tot tijd neer op een takje of gewoon op de grond. Een enkele maal echter vloog een wesp zoekend van plant naar plant zonder ergens te landen. Ze bleef alleen maar even stil in de lucht hangen, voor een takje, en vloog dan weer naar een volgend takje om ook hier weer even voor te blijven hangen. Een paar maal zag ik een van de wespen in actie met een bij. Als de wesp toevallig op minder dan een halve meter van haar vandaan een bij zag, draaide ze zich abrupt om, zodat ze tegenover haar prooi kwam, vloog er dan op af en hield enkele centimeters voor de bij haar vlucht in om bewegingloos, als een zweefvlieg, in de lucht te blijven hangen. Met een plotselinge duik wierp ze zich vervolgens op de bij. En vanaf dat ogenblik gebeurde alles precies zoals ik het reeds in mijn jampot-experimenten had gezien. Er had een korte worsteling plaats, waarbij beide partijen heftig zoemend en als dollen
rondtollend dwars door de heidestruiken omlaag vielen. Het volgende moment had de wesp de bij in een gunstige positie gemanoeuvreerd en stak zij haar slachtoffer. Dan drukte ze de bij stevig tegen zich aan, likte de nectar op en vloog na een minuut of wat in noordwestelijke richting weg, terug naar de kolonie.
Dit was dus de hele gedragsketen. In de loop van de volgende dagen kreeg ik verschillende zaken te zien die mijn begrip nog vergrootten. De eerste reactie van de wesp, het zich omdraaien tijdens de vlucht of als ze ergens zat, en daarna het zich recht tegenover de bij plaatsen, hield geen verband met de heersende windrichting. Deze reactie kon daarom ook onmogelijk afhangen van het reukvermo- | |
| |
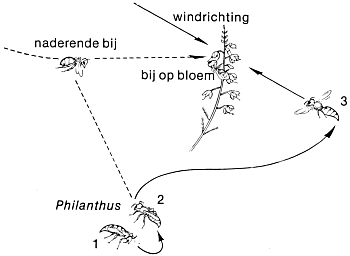
Alvorens een vliegend insekt aan te vallen, zal Philanthus het altijd beneden de wind naderen om even de geur ervan te controleren.
gen, in tegenstelling tot het gedrag dat de wesp vertoonde bij het opsporen van een zoekgeraakte bij. Ik veronderstelde dat de prikkel van visuele aard moest zijn, vooral omdat de maximale afstand waarop een wesp een bij bleek te kunnen ontdekken, minder was dan een halve meter, wat goed overeen leek te komen met de groote van hun gezichtsveld, die waarschijnlijk in de buurt van 1 o lag. Dit zou betekenen dat een voorwerp ter grootte van een bij onmogelijk gezien kon worden op een afstand van meer dan dertig centimeter. Maar hoe was het dan mogelijk dat de wespen onderscheid maakten tussen een honingbij en andere insekten van ongeveer gelijke afmeting? Ik zag weldra dat zij dit bij deze afstand ook niet konden. Zij reageerden ook op andere insekten, volgen zelfs op hen af; ik zag reacties op hommels, vliegen en kleine torretjes. Het gedrag jegens deze insekten maakte mij nog iets anders duidelijk ook. Zij benaderden hen altijd op dezelfde manier als honingbijen en bleven vlak voor hen stil in de lucht hangen, maar vlogen dan verder. Dit ‘bidden’ vlakbij de potentiële prooi gebeurde altijd met de neus in de wind, met welk insekt zij ook geconfronteerd werden. Het leek erop dat zij de geur controleerden.
Nu zou het niet moeilijk meer zijn om dit experimenteel na te gaan, als ik maar voldoende wespen zover kon krijgen om een voldoende
| |
| |
groot aantal experimenten te doen. Dit bleek heel goed mogelijk. Ik doodde een paar bijen en bond ze vast aan een dunne draad die ik tussen twee takjes spande, die verticaal, een halve meter uit elkaar waren geplant. Ik had mijn opstelling zo gemaakt dat de bijen precies voor en in de luwte van een paar Calluna-struikjes hingen. Daarna nam ik mijn post weer in en wachtte af. Diezelfde dag nog kwamen er verschillende wespen langs, jagend van plant naar plant. De eerste twee passeerden zonder ook maar enige andacht te schenken aan mijn namaakbijen en, ongeduldig als ik was, begon ik al te wanhopen. Maar juist toen de derde wesp mijn lokaas was genaderd, deed een briesje de dode bijen bewegen en meteen kreeg de wesp hen in het vizier, bleef voor een van hen in de lucht hangen en wierp zich er toen op. Ze stak het dier, daarbij ondersteboven hangend en met een poot om de draad geklemd, greep haar slachtoffer, drukte het tegen zich aan en likte de nectar op. Toen probeerde ze weg te komen, wat haar rond deed tollen als een acrobaat aan de vliegende trapeze. Luid zoemend bleef ze proberen met haar buit er vandoor te gaan, maar tenslotte gaf ze het op.
Nu wist ik hoe ik mijn volgende experiment moest opzetten. Allereerst bevestigde ik een extra draad aan een van de takken. Door hier zachtjes aan te trekken kon ik de hele opstelling op en neer laten schommelen en daardoor ontdekte ik dat mijn eerste waarneming geen puur toeval geweest was: de flauwe bewegingen trokken de aandacht van de wespen die dicht genoeg in de buurt waren om de bijen te zien. Vervolgens bood ik in plaats van uitsluitend dode bijen verschillende andere imitaties aan. Mijn basisopstelling bestond uit een pas gedode bij, een droog, dood Calluna-takje, ongeveer zo groot als een bij, een dode bij die ik eerst even in alcohol en later in ether had bewaard en daarna gedroogd had om alle geur ervan af te halen, verder een stokje dat samen met levende bijen was geschud zodat het naar bijen was gaan ruiken, en tenslotte een bij die ik geurloos had gemaakt en vervolgens opnieuw een geur had gegeven door hem met levende bijen te schudden. De levende bijen zullen het wel niet prettig hebben gevonden, zo geschud te worden, maar voor zover ik kon nagaan liepen ze er geen ernstig letsel bij op en ik voelde dan ook geen wroeging als ik ze op die manier dwong mij een handje te helpen. In de loop van dat seizoen behaalde ik enkele zeer bevredigende resultaten. Er zat hoegenaamd geen variatie in de reacties van de ongeveer honderd wespen die mijn proefopstelling bezochten. Alle namaakprooien werden even dikwijls benaderd en
| |
| |
gefixeerd. Alle geurende prooien werden eveneens ‘gevangen’. Het geurende stokje echter kreeg geen enkele steek, terwijl de bijen die ik opnieuw van een geur had voorzien wel steken opliepen. Deze resultaten maakten de hele geschiedenis duidelijk. De eerste reactie was inderdaad van visuele aard. Tijdens het bidden in de lucht controleerde de wesp de geur. Nadat de prooi gevangen was, bepaalden nieuwe prikkels of hij al dan niet gestoken zou worden en daarvoor kwamen ofwel bepaalde visuele stimuli ofwel tactiele prikkels in aanmerking. De aard van de eerste visuele prikkelsituatie was betrekkelijk eenvoudig: ieder bewegend object dat ongeveer de grootte van een honingbij bezat werd aan een nader onderzoek onderworpen. De afmetingen konden binnen zeer ruime grenzen variëren: zelfs zeer grote hommels werden geïnspecteerd, evenals kleine vliegjes die maar half zo groot waren als een honingbij. Indachtig het feit dat de wespen in staat waren om een bij op een afstand van enkele meters te ruiken, verbaasde ik mij erover dat geen enkele jagende wesp ooit mijn geurende namaaksels van een grotere afstand dan een halve meter waarnam. Ik plaatste zelfs tien bijen in een open buisje dat ik in de heide verborg; hoewel hun geur zo sterk was dat zelfs ik hen gemakkelijk op een halve meter afstand kon ruiken, was er geen wesp te bekennen die erop reageerde. Het verschil met het gedrag van een wesp die haar zoekgeraakte bij opspoort, was werkelijk verbazingwekkend. Klaarblijkelijk waren al deze jagende wespen ingesteld op een visuele prikkel, en werden ze pas als ze deze hadden ontvangen ‘geprogrammeerd’ om te reageren op geurprikkels. Dit maakte een diepe indruk op mij; de resultaten waren zo ontzettend duidelijk. Toch vond ik het
allemaal maar onbegrijpelijk. Later leerde ik dat dit een heel normaal kenmerk is van een keten van opeenvolgende gedragspatronen, wat natuurlijk het begrip ervan niet gemakkelijker maakt, maar wat ons wel toont dat wij te maken hebben met een algemeen verschijnsel.
Deze zelfde buizentest konden wij herhalen met levende bijen, zodat behalve de geur nu ook het gezoem op enige afstand kon worden waargenomen. Ik deed dit experiment een paar maal, maar weer reageerde er geen enkele wesp en ik denk ook eigenlijk niet dat wespen kunnen horen.
Voor het eind van het seizoen voltooide ik nog één ander onderdeel van mijn onderzoek: als de wesp de prooi op zijn geur had getoetst en besloten had hem te bespringen, welke prikkels begeleidden dan deze sprong? De pijlsnelle beweging en de feilloze precisie ervan verschilden hemelsbreed van de aarzelende, onhandige manieren
| |
| |
van de wesp die op zoek was naar een verloren prooi en ik kon mij moeilijk voorstellen dat deze sprong een reactie was op een geurprikkel; het stond vrijwel vast dat de wesp daarbij op haar ogen afging. Nu maakte ik een opstelling die iets ingewikkelder was voor de wespen. Ik bood hen een geurend stokje aan, dat enkele centimeters achter een niet-geurend stokje hing, aan de zijde waarvan de wind vandaan kwam. Zoals gewoonlijk kwamen de wespen op de in de lucht hangende stokjes af en sprongen er naar. Ze grepen echter nooit het geurende stokje, maar altijd het andere, dat het dichtst bij hen hing. Dit kon niets anders betekenen dan dat de geur weliswaar de sprong ontketende, maar dat deze door een visuele prikkel, nl. het zien van de prooi, geleid werd.
| |
Mogelijkheden voor verder onderzoek
De resultaten van deze experimenten verschaften ons meer dan voldoende stof tot nadenken. Bijvoorbeeld het gegeven dat wespen altijd op een afstand van zeven tot tien centimeter van de prooi doodstil in de lucht blijven hangen, en dat vrijwel zonder variatie, toonde dat zij op de een of andere manier in staat zijn om vrij nauwkeurig afstanden te schatten. Bovendien moesten ze op zijn minst een verschil in grootte kunnen zien tussen bijvoorbeeld een honingbij en een grote hommel. Toch leerden zij nooit onderscheid te maken tussen een ongewone prooi of een namaaksel, louter op het gezicht, want ze bleven reageren op mijn namaakbijen, zelfs op de abnormaal grote die ik hen nu en dan presenteerde. Deze hardleersheid vormde een schril contrast met de werkelijk verbijsterende leerprestaties op het gebied van bakens-herkennen, die wij ontdekten tijdens onze experimenten betreffende hun oriëntatievermogen. Wij konden niet anders dan aannemen dat de wespen op bepaalde tijden erop ingesteld waren iets te leren, en op andere tijden niet. Op die wijze werden wij opnieuw geconfronteerd met een zekere starheid in gedrag, met zekere beperkingen aan hetgeen waartoe de wespen in staat waren of dat zij op een bepaald moment konden leren. In andere opzichten constateerden wij echter weer een ongelooflijke soepelheid. Er was bijvoorbeeld een bijna onbeperkte variatie in de bewegingen die de wespen maakten om een bij zo te draaien dat hij in de juiste positie lag om gestoken te worden; dit zou vanzelfsprekend ook niet anders kunnen, want zij moesten tegenspel bieden aan de heftige en onvoorspelbare bewegingen van de bij.
| |
| |
Vanzelfsprekend is het juist dit ontbreken van flexibiliteit in zoveel opzichten van het gedrag dat ons vertrouwen in de uiteindelijke effectiviteit van dit type onderzoek in stand hield. De basis voor dit vertrouwen lijkt te worden gevormd door het feit dat de beperkingen van het gedrag van een dier zich steeds duidelijker manifesteren naarmate wij er meer over ontdekken. Deze beperkingen zijn misschien niet altijd even begrijpelijk, ze zijn niettemin reëel. Om twee voorbeelden te noemen: een wesp zal nooit zand wegharken door het naar voren te duwen of zijn kaken te gebruiken, hoewel dat gezien zijn spierstelsel heel goed mogelijk zou zijn. Wanneer het dier echter een kiezelsteentje moet verplaatsen, zal het altijd proberen dit met zijn kaken vast te pakken en het nooit met zijn lichaam gaan wegduwen.
Vijf zomers hadden wij nodig gehad om dit beeld van het leven van een bijenwolf te construeren, ontegenzeggelijk een lange periode, maar dit type werk vordert nu eenmaal nooit snel; het wordt vertraagd door ongunstige weersomstandigheden, doordat men in het wild levende dieren nu eenmaal niet naar zijn hand kan zetten en door nog allerlei andere problemen die een wezenlijk onderdeel vormen van veldwerk. Toch zou het absoluut onmogelijk zijn geweest, dit onderzoek in een laboratorium te doen; het was een kwestie van in het veld experimenteren of helemaal niet experimenteren. En er waren ook vele voordelen aan verbonden. Een daarvan was de mogelijkheid om het hele gedrag van een soort waar te nemen. Daardoor kon Van Beusekom zijn onderzoek uitbreiden tot het voedingsgedrag van de wesp, de reacties op bloemen, hun geur en kleur. Dit alleen al biedt genoeg materiaal voor een boeiend artikel, maar ik zal daar hier verder niet op ingaan.
Omstreeks het einde van de jaren dertig begonnen de aantallen wespen af te nemen. Geleidelijk aan was het ons duidelijk geworden dat Philanthus, doorgaans zeldzaam in West-Europa, gedurende de late jaren twintig een geweldige bloeiperiode had doorgemaakt. Tijdens de topjaren, rond 1930, was onze aandacht op hen gevallen en juist hun massale voorkomen had ons tot dit experimentele onderzoek gestimuleerd. Toen de wespen na 1935 in aantal begonnen af te nemen, bereikten wij spoedig het stadium waarop experimenten op grote schaal niet langer mogelijk waren. Daarom schakelden wij over op andere dieren. Tegen die tijd echter hadden wij zoveel mogelijkheden voor ander onderzoek ontdekt, dat ik besloot om Hulshorst te blijven gebruiken als studieterrein voor mijn studenten. Wij hebben deze beslissing nooit hoeven te betreuren. |
|
