|
| |
| |
| |
Han Collewijn
Kleurenzien: een fysiologische blik
Kleuronderscheid speelt in het leven van de mens vanouds een grote rol. Hierbij moeten we niet in de eerste plaats denken aan ‘luxe’ hedendaagse aspecten zoals de esthetische beleving van een rijkdom aan kleuren en hun combinaties die we vaak ondergaan, maar vooral aan heel elementaire, voor het overleven essentiële functies zoals het snel onderscheiden en herkennen van beschikbaar voedsel, bij voorbeeld eetbare vruchten. Kijk maar naar Fig. 1. In de rechter plaatjes is de kleur uit de linker plaatjes verwijderd, en is er alleen licht/donker. Zonder kleurinformatie vallen de kersen te midden van de bladeren nauwelijks op, en is het onmogelijk om de rijpe van de onrijpe tomaat te onderscheiden. Ook in goed gecontroleerde metingen aan het waarnemingsvermogen van proefpersonen blijkt ons visuele systeem bij uitstek gevoelig te zijn voor zulke kleurnuances. Dit onderscheidingsvermogen vereist een complexe organisatie van ogen en hersenen. Dat die in de loop van de evolutie ontwikkeld is en standgehouden heeft, geeft aan dat dit vermogen veel bijdraagt aan onze overlevingskansen. Dit artikel wil de fysiologische grondslagen van het kleurenzien belichten, rekening houdend met moderne wetenschappelijke inzichten, maar zonder in te gaan op technische details.
| |
Ogen en hersenen
Een summier overzicht van het menselijke visuele systeem is geschetst in Fig. 2. Elk oog vormt via een optisch systeem een afbeelding van de omgeving op zijn achterwand, de retina. Dit proces kan in eerste instantie vergeleken worden met dat van een camera; de retina bevat de lichtgevoelige elementen. Met de cameralens komen overeen de gebogen oppervlakken van hoornvlies (de voorkant van het oog) en de inwendige lens; veranderlijke kromming van de ooglens zorgt voor scherpstelling (accommodatie) en de iris vormt een diafragma (de veranderlijke pupilgrootte). Elk oog overziet een hoek van ongeveer 150 graden van de omgeving (zoals een groothoeklens), namelijk 90 graden naar opzij en 60 graden naar de kant van de neus. In het midden overlappen de twee gezichtsvelden zodat we een grote hoek (ongeveer 120 graden) met beide ogen zien. In Fig. 2 wordt de appel dus op de retina's van beide ogen af-
| |
| |

1. Voor wie normaal kleuren ziet zijn de kersen (a) en de rijpe tomaat (c) moeiteloos te herkennen. Zonder kleurinformatie is dit lastig (B) of zelfs onmogelijk (d). Voor iemand met rood-groen blindheid is het kleuronderscheid in a en c ook niet te maken, de twee kleuren zien er het zelfde uit.
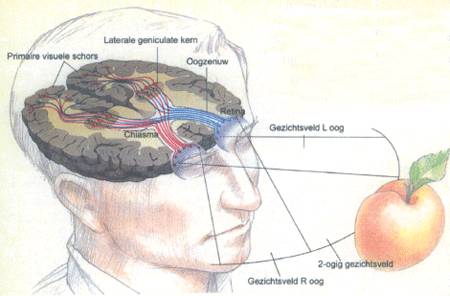
2. De verbindingen van de retina van het linker oog (in blauw) en het rechter oog (in rood) met de visuele hersenschors. (Naar: Scientific American, september 1992, p. 36).
| |
| |
gebeeld, overigens gezien vanuit twee standpunten (gescheiden door de afstand tussen de ogen). Dit laatste vormt de basis voor stereoscopisch dieptezien, waar ik hier niet op in ga. De retina bevat een mozaïek van lichtgevoelige cellen, de fotoreceptoren, en bovendien een groot aantal zenuwcellen, die een eerste signaalbewerking uitvoeren. De retina is eigenlijk een stukje zenuwstelsel en wordt in de embryonale ontwikkeling dan ook gevormd door een uitgroeisel van de hersenaanleg.
Een belangrijk verschil tussen het oog en een goed fototoestel is verder dat de kwaliteit van de optische afbeelding en ook van de retina alleen in een klein centraal deel hoog is. Alleen dit centrale deel van de retina, de fovea, heeft voldoende oplossend vermogen - gezichtsscherpte - om fijne details te onderscheiden, zoals klein gedrukte tekst. Daar de fovea slechts één tot enkele graden gezichtshoek bestrijkt, moeten onze ogen voortdurend bewegingen uitvoeren om een groter gebied in detail af te tasten.
Uitlopers van zenuwcellen in de retina verlopen via de oogzenuw naar de schedelholte, kruisen voor de helft in het chiasma en eindigen voornamelijk in de laterale geniculate kern van de hersenen. Daar vindt overschakeling op andere zenuwcellen plaats, waarvan de uitlopers eindigen in de primaire visuele hersenschors, die achterin de hersenen ligt (occipitaal), grotendeels verborgen in het middenvlak waar de linker en rechter hersenhelft tegen elkaar liggen. Door de gedeeltelijke kruising in het chiasma ontvangt de rechter hersenschors informatie over de linker helft van het gezichtsveld van beide ogen, en vice versa. Op deze wijze kan de schors de informatie van de twee ogen integreren. Het oppervlak van de retina, en dus de (afbeelding van de) omgeving wordt op een topografische, maar vervormde wijze geprojecteerd in de hersenschors, waarbij het centrale gezichtsveld de grootste representatie heeft. De organisatie van de primaire visuele schors is zeer complex, en van daar uit wordt informatie verder geschakeld naar een groot aantal aansluitende schorsgebieden die allemaal betrokken zijn bij het totstandkomen van de uiteindelijke visuele gewaarwording, de herkenning van objecten, de ruimtelijke oriëntatie en de integratie van visuele met andere informatie.
| |
Licht
Onder ‘licht’ verstaan we het deel van het elektromagnetische spectrum dat met het oog waarneembaar is. Het bestaat uit lichtkwanten (de kleinst bestaanbare hoeveelheden licht), de fotonen. Een foton heeft zowel een deeltjes- als een golf-karakter. Elk foton heeft zijn karakteristieke, onveranderlijke golflengte (die omgekeerd evenredig is met zijn energie) en beweegt zich in een homogeen medium (zoals lucht of water) rechtlijnig voort met de lichtsnelheid (in het luchtledige bijna 300.000 km/s). Voor
| |
| |
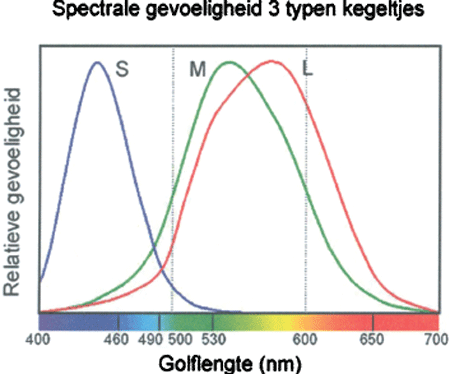
3. De gevoeligheid van de 3 typen kegeltjes voor de verschillende golflengten van het zichtbare licht. Op de horizontale balk het spectrum dat zichtbaar wordt als een straal zonlicht via een prisma op een wit scherm valt.
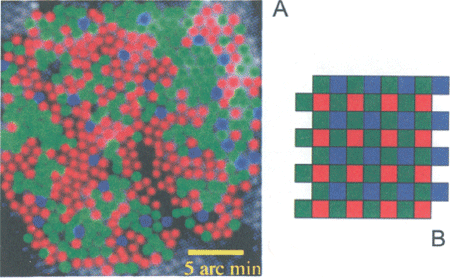
4. a. De verdeling van l, m en s kegeltjes (resp. rood, groen en blauw) in een levend menselijk oog, zichtbaar gemaakt met een adaptieve optische techniek.
(Ontleend aan: Roorda & Williams 1999, Nature, 397: 520-522).
b. Ter vergelijking het rgb mozaïek van de lichtsensor van een digitale camera (niet op schaal t.o.v. a).
| |
| |
de mens zichtbaar licht bestrijkt het golflengtegebied van ongeveer 400 tot 700 nanometer (1 nanometer - 1 nm - is een miljardste meter). De ligging van dit spectrale gebied is geen toeval. De zonnestraling die het aardoppervlak bereikt bevat in dit gebied een grote hoeveelheid energie, die een aaneengesloten verdeling vertoont, dat wil zeggen dat fotonen van alle mogelijke golflengtes binnen dit gebied rijkelijk aanwezig zijn (het spectrum is continu).
Verder vertonen fotonen in dit gebied een voor het zien essentiële interactie met onze materiële omgeving: ze worden deels weerkaatst en deels geabsorbeerd, afhankelijk van hun golflengte en de eigenschappen van een voorwerp. Ten slotte zijn water en waterige substanties (zoals de optische media van ons oog) voor het zichtbare deel van het spectrum transparant, maar doet zich op een lucht-water grensvlak refractie (richtingsverandering) van de fotonen voor, wat weer een voorwaarde is voor het vormen van een afbeelding via gebogen grensvlakken, zoals ons bolle hoornvlies. Een zelfde refractie doet zich voor op een overgang van lucht naar glas, en dit is de basis voor de werking van lenzen en prisma's.
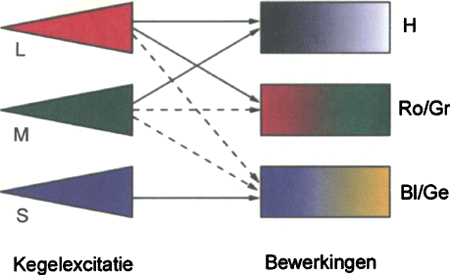
5. De bewerking van de 3 kegelsignalen tot 3 informatiestromen naar de hersenen: Helderheid (H), Rood/Groen balans (Ro/Gr) en Blauw/Geel balans (Bl/Ge). (Naar: Boynton, 2002, Current Biology, 12: R838-R840).
Het verband tussen golflengte en kleur werd voor het eerst duidelijk gedocumenteerd door de beroemde experimenten van Newton (1704), waarbij hij een bundel zonlicht door een prisma liet vallen en constateerde dat het ‘witte’ licht werd gesplitst in een reeks van ‘gekleurde’ banden, verlopende van ‘rood’ via ‘oranje’, ‘geel’ en ‘groen’ naar ‘blauw’. Dit verloop - dat we ook zien in de regenboog - is weergegeven in de ho- | |
| |
rizontale balk in Fig. 3. De aanhalingstekens staan er omdat we op dit punt een scherp onderscheid moeten maken tussen de fysische en psychische eigenschappen van het licht. Fysisch worden fotonen gekarakteriseerd door hun golflengte; de golflengtes worden van elkaar gescheiden door een prisma omdat de richting van korte golflengtes sterker wordt afgebogen dan die van langere golflengtes. Golflengtes hebben op zichzelf geen ‘kleur’, zoals Newton al benadrukte in de bespreking van zijn experimenten, die overigens een eeuw later de uitzinnige woede zouden opwekken van de dichter Goethe, die zijn eigen - totaal subjectieve - kleurentheorie als zijn absolute meesterwerk beschouwde.
Dat verschillende golflengtes leiden tot verschillende kleurgewaarwordingen wordt mogelijk doordat de retina mechanismen bevat om golflengtes van elkaar te onderscheiden, waarna neurale processen deze uiteindelijk koppelen aan een kleurgewaarwording, die zuiver psychisch is. Een rijpe tomaat weerkaatst vooral licht met langere golflengtes (boven 500 nm) en absorbeert vooral de kortere golflengtes (beneden 500 nm). Alleen het weerkaatste licht bereikt ons oog; hierin overheersen in dit geval de lange golflengtes en deze verdeling activeert uiteindelijk de gewaarwording ‘rood’.
| |
Fotoreceptoren
De retina bevat twee hoofdgroepen lichtgevoelige cellen (fotoreceptoren), staafjes en kegeltjes. De staafjes zijn zeer talrijk, maar ontbreken in de centrale fovea. Ze zijn zeer gevoelig, en worden onder optimale omstandigheden zelfs geactiveerd door één enkel foton. Ze zijn actief bij het zien bij zeer lage verlichtingssterktes, zoals bij nacht zonder straatverlichting en met alleen een sikkeltje maan. Alle staafjes hebben de zelfde spectrale gevoeligheid en ze maken dus geen onderscheid tussen golflengtes. Verder werken de staafjes in grote groepen samen. Het gevolg is dat we bij schaars licht alleen een grof beeld zien zonder detail of kleur, zoals direct blijkt als we in het bijna-duister proberen te lezen of kleuren te onderscheiden.
Kleuronderscheid is alleen mogelijk bij voldoende licht, waarbij een ander type fotoreceptoren, de kegeltjes, actief is. De retina van de mens bevat 3 typen kegeltjes, die hun grootste gevoeligheid hebben voor licht met relatief lange golflengtes (l), golflengtes in het middengebied (m) en korte golflengtes (s; voor ‘short’). De spectrale gevoeligheidscurven van deze 3 typen kegeltjes zijn (bij benadering, en met een arbitraire verticale schaal) weergegeven in Fig. 3. Hun maximale gevoeligheden liggen bij resp. 565, 535 en 440 nm, maar ze vertonen elk een breed gevoeligheidsgebied. Voor een m-kegeltje betekent dit dat fotonen met golflengte 535 nm de grootste kans hebben om geabsorbeerd te worden, terwijl fotonen
| |
| |
met golflengte 600 nm een half zo grote kans op absorptie hebben. Elke absorptie van fotonen, van welke golflengte ook binnen het gevoelige gebied, zal het kegeltje echter activeren (dit is het ‘univariantie principe’). Voor de andere 2 typen kegeltjes geldt hetzelfde, met verschuiving van het gevoelige gebied. De spectrale gevoeligheden zijn niet alleen breed, maar vertonen ook veel overlap, vooral tussen de l en m kegeltjes. Uit deze verdeling volgt dat de activering van één kegeltype geen uitsluitsel geeft over de golflengte van het activerende foton; het gevoelige gebied is daarvoor veel te breed. Alleen de vergelijking van de activiteiten van de verschillende kegeltjestypen kan hier uitkomst bieden. Zo zal een stroom van fotonen van 600 nm volgens de curves van Fig. 3 een bijna tweemaal zo grote kans hebben om door een l-kegeltje geabsorbeerd te worden dan door een m-kegeltje. Dus als de l-kegeltjes tweemaal zo actief zijn als de m-kegeltjes, dan zou dit in principe wijzen op de aanwezigheid van veel fotonen rond de 600 nm.
Een bijzonder interessant punt is hoe de 3 typen kegeltjes ruimtelijk verdeeld zijn over het retina oppervlak. Met anatomische technieken was hier tot voor kort slechts beperkte informatie over te krijgen, alleen de s-kegels waren te onderscheiden van de groep van m- en l-kegels, die onderling geen anatomische verschillen vertonen. In de late 90er jaren slaagden Roorda en Williams (in Rochester, ny) er toch in het kegelmozaïek direct zichtbaar te maken, en wel in het levende menselijke oog, door het gebruik van een zeer geavanceerde oogspiegeltechniek waarin ‘adaptieve optica’ werd toegepast, dezelfde techniek die gebruikt wordt in de modernste astronomische licht-telescopen. Hierbij worden onvolmaaktheden in de optische media gemeten en vervolgens gecompenseerd via een spiegeloppervlak dat op een groot aantal plaatsen mechanisch vervormd kan worden. Dit leidde tot een spectaculaire verscherping van de afbeelding, waarbij de kegels individueel zichtbaar werden. Het onderscheid tussen de 3 kegeltypen kon gemaakt worden door het oog eerst sterk te belichten met een smal golflengtegebied in het l, m of s gebied, waarbij het meest geactiveerde type kegeltje een (tijdelijke) verandering ondergaat door ‘bleking’ van het fotopigment, die zichtbaar is. Een resultaat voor een retinagebiedje net buiten de fovea is gereproduceerd in Fig. 4 a. De l, m en s kegeltjes zijn hierin ter illustratie rood, groen en blauw gekleurd; in werkelijkheid hebben deze kegeltjes natuurlijk niet deze kleuren, en is het hoe dan ook te eenvoudig om van rode, groene en blauwe kegeltjes te spreken (de l-kegeltjes zijn bovendien maximaal gevoelig in het ‘gele’, niet in het ‘rode’ gebied, zie Fig. 3). Belangrijke conclusies uit dit onderzoek waren:
(1) het mozaïek van de 3 typen kegeltjes is zeer onregelmatig; (2) de s-kegeltjes zijn het geringst in aantal en liggen vrij ver van elkaar; (3) de m- en l-kegels liggen in clusters, waarbij l- en
| |
| |
m-clusters geen regelmatig patroon vormen; (4) de verhouding tussen de aantallen l- en m-kegels varieert sterk tussen individuen, zonder dat dit leidt tot duidelijke verschillen in hun kleurenzien. Verder is bekend dat de centrale fovea alleen l- en m-kegeltjes bevat, dus geen s-kegels (en ook geen staafjes).
| |
Bewerkingen: drie informatie stromen
De activiteitsniveaus van de kegeltjes worden al in de retina zelf, door circuits van verschillende typen neuronen, bewerkt tot een drietal gescheiden informatiestromen (Fig. 5) die via de oogzenuw en de laterale geniculate kern naar de hersenen lopen. Men kan spreken van drie ‘kanalen’. Eén daarvan bepaalt de balans in activering van l- en m-kegels (l vs.m) en vormt het rood/groen kanaal. (Het is aannemelijk dat de neurale circuits zich aanpassen aan de locale en individuele verschillen in aantallen l en m kegeltjes). Een tweede bepaalt de balans tussen enerzijds de activering van de s-kegels en anderzijds de gezamenlijke activiteit van de l en m kegels. Dit is het blauw/geel kanaal. Deze twee kanalen vormen ‘opponente’ systemen, waarvan de componenten binnen één kanaal elkaar uitsluiten: we zien een kleur nooit als groenig rood of gelig blauw. Wel bestaan geel-groen, rood-geel, blauw-groen en blauw-rood (paars). Naast deze kleurverhoudingen, die de tint bepalen, wordt in een derde kanaal de helderheid (de lichtheid) bepaald door de gezamenlijke activiteit te bepalen van de l en m kegels (l+m). De blauwe kegels dragen niet bij aan deze helderheid, alleen aan het onderscheid blauw/geel. Dit helderheidskanaal codeert dus voor donker (weinig activiteit) tot licht (veel activiteit), op een wijze zoals de zwart-wit versies in Fig. 1, waarbij de tussenwaarden tussen de lichtste en donkerste partijen ‘grijs’ genoemd worden.
De waarden van de drie kanalen worden per plek in de retina berekend, om aldus een totaalbeeld van de verschillende kleuren en helderheden op te bouwen. Het ruimtelijke detail hiervan is, zoals eerder vermeld, aanmerkelijk groter in het centrum van het gezichtsveld dan in de periferie.
In de ruimtelijke verwerking speelt verder het contrast tussen aangrenzende gebieden een grote rol. Zulke contrasten worden versterkt gezien, als de overgang vrij scherp is. Een rood vlak lijkt intenser rood als het omgeven wordt door een groot groen vlak. Een helder vlak wordt extra helder gezien als het omgeven wordt door een donker vlak. Zelfs wordt een neutraal gekleurd (wit of grijs) vlak bijv. groenig gezien als het omgeven wordt door intens rood, en vice versa (hetzelfde geldt voor geel en blauw). Deze effecten van simultaan contrast dragen bij aan het scherper waarnemen van betrekkelijk kleine verschillen in kleur of helderheid. (Zeer geleidelijke overgangen, bijv. een gradueel verloop van licht naar
| |
| |
donker grijs, worden daarentegen nauwelijks opgemerkt).
Deze bewerkingen worden al in de retina en de laterale geniculate kern uitgevoerd en voortgezet in de hersenschors. Omstreden is nog in hoeverre de drie stromen in de hersenschors gescheiden blijven dan wel geïntegreerd worden. Rond 10-15 jaar geleden leken er goede aanwijzingen te zijn dat de kanalen verregaand gescheiden bleven, en dat er bijvoorbeeld een duidelijk afgegrensd hersenschorsgebied bestond dat bij uitstek de kleurgewaarwording verzorgde. Meer recent onderzoek brengt veel twijfel aan deze scherpe lokalisatie van functies en ondersteunt meer een integrale verwerking van alle aspecten van de visuele waarneming in sterk onderling verbonden schorsgebieden.
| |
Trichromatie, kleurenweergave
Het feit dat de mens talloos veel kleuren ziet op grond van slechts drie typen kegeltjes, dus een trichromaat is, heeft grote praktische consequenties. Het betekent namelijk dat iedere waarneembare kleur in principe nagemaakt kan worden door een mengsel van drie lichtbronnen van geschikt gekozen golflengtes, bijv. rond de 650, 530 en 430 nm (het rode, groene en blauwe gebied) die door het instellen van de juiste onderlinge intensiteits-verhoudingen dezelfde activering van de drie kegeltjestypen bewerkstelligen als de originele kleur. Zo kan de kleur van een testlicht waarin alleen fotonen van 570 nm voorkomen (monochromatisch geel) voor het oog gemakkelijk nagebootst worden door een adequaat mengsel van lichtbronnen rond 650 nm (rood) en 530 nm (groen); het blauwe licht blijft hierbij op intensiteit nul staan. Fysisch zijn testlicht en mengsel totaal verschillend, in onze waarneming zijn ze volstrekt gelijk. Voor het blauwe en blauw-groene gebied zijn alle drie kleuren nodig. Dat wij trichromaten zijn vergemakkelijkt de reproductie van kleuren aanmerkelijk, want zodoende kan het beeldscherm van de televisie en computer (bij benadering) alle benodigde kleuren genereren door het in de juiste mengverhouding laten oplichten van de grondkleuren rood, groen en blauw. Ook digitale kleurenfotografie gebruikt dit principe: Fig. 4 b toont een stukje van de lichtgevoelige sensor van een digitale camera, die - anders dan de retina - volgens een volmaakt regelmatig mozaïek van drie kleurgevoelige gebieden, rood, groen en blauw, geconstrueerd is (het rgb systeem). Ook kleurendruk geschiedt met een beperkt aantal kleurpigmenten; we gaan hier even voorbij aan de verschillen tussen mengen van licht (additief) en van pigmenten (subtractief).
Wanneer de drie kegeltypes in dezelfde verhouding geactiveerd worden als door het zonlicht, wordt het als kleurloos, ‘wit’ ervaren. Een ‘wit’ voorwerp reflecteert alle golflengtes van het erop vallende licht in gelijke mate. In dat geval geven de twee kleur-opponente kanalen een ‘neutraal’
| |
| |
signaal (niet rood en niet groen, niet blauw en niet geel), en is vooral het helderheidskanaal van belang: wit versus zwart.
Overigens past het systeem zich goed aan als de samenstelling van het licht afwijkt van die van het natuurlijke daglicht. Het licht van een gloeilamp bevat veel meer lange golflengtes dan zonlicht; toch blijven we de omgeving globaal in dezelfde kleuren zien. Dit verschijnsel heet kleur-constantheid. Over de mechanismen hiervan zijn vele theorieën, maar de constantheid heeft wel zijn grenzen. Bij het kopen van kleding lopen we toch liefst even naar het daglicht om de kleur goed te beoordelen. Kleuren kunnen niet onderscheiden worden als de verlichting slechts in een nauw golflengtegebied plaatsvindt (monochromatisch licht). Een goed voorbeeld hiervan is het licht van de lage druk natrium lamp, die vaak gebruikt wordt voor de verlichting van buitenwegen. Bij dit vrijwel monochromatische (580 nm; geel-oranje) licht is kleuronderscheid onmogelijk. Tenslotte is nog van belang het begrip verzadiging. Als gekleurd licht gemengd wordt met ‘wit’ licht, wordt de kleur minder intens, bleker, meer pastelachtig. Dit noemen we een onverzadigde kleur. Felle kleuren bevatten geen neutrale, witte component, ze zijn verzadigd.
| |
Evolutie en genetica
De fotoreceptoren bevatten een lichtgevoelig pigment. Het molecuul hiervan bestaat uit twee delen: retinal (afgeleid van vitamine a) en opsine, een eiwit. Als dit complexe molecuul een foton absorbeert, verandert het retinal-deel van vorm; dit leidt tot een cascade van biochemische reacties in de receptorcel met als gevolg activering. (Voor de activering van een staafje is minimaal één foton nodig, voor die van een kegeltje enkele honderden). Het retinal-deel is voor alle fotoreceptoren gelijk. De verschillen zitten in het eiwitdeel, het opsine, dat bij de mens in vier vormen voorkomt: één voor de staafjes en drie andere voor de drie kegeltypen. Dit opsine-deel bepaalt de vorm van het absorptiespectrum, en daarmee de spectrale gevoeligheid van de verschillende fotoreceptoren. Een eiwit is opgebouwd uit een reeks aminozuren; de volgorde hiervan wordt gecodeerd door een gen. Er zijn dan ook afzonderlijke genen voor de verschillende typen opsinen (en dus fotoreceptoren), en dit geeft de sleutel voor de studie van genetische afwijkingen in het kleurenzien, voor het bepalen van de verschillen tussen dieren, en voor het volgen van de ontwikkeling van het kleurenzien in de evolutie. Dit moleculair-biologische onderzoek heeft de laatste jaren een grote vlucht genomen. De details zijn buitengewoon ingewikkeld, maar ik wil hier enkele interessante punten noemen.
In de evolutie was er oorspronkelijk slechts één type fotoreceptor met één fotopigment. Met dit monochromatische zien kon uitsluitend helder- | |
| |
heid worden waargenomen (zwart/wit). Door duplicatie en divergentie van het betrokken gen zijn subtypes ontstaan met verschillende opsines, en dus een verschillende spectrale gevoeligheid. De oudste differentiatie is die tussen langegolf-gevoelige fotopigmenten (groter dan 500 nm; l) en kortegolf-gevoelige (kleiner dan 500 nm; s). Analyse van genetische drift suggereert dat deze differentiatie al een miljard jaar geleden heeft plaatsgevonden, lang voor het ontstaan van gewervelden (ongeveer 495 miljoen jaar geleden) of zoogdieren (ongeveer 220 miljoen jaar geleden). Onze huidige l en s kegeltjes zijn natuurlijk niet identiek met deze oude vormen, maar hun langdurige gescheiden evolutie wordt bevestigd door de grote onderlinge verschillen in de opbouw van hun opsines: de s en l opsines verschillen in ongeveer 200 van de totaal ongeveer 400 aminozuren.
Een systeem met alleen s en l kegeltjes is dichromaat, en kan twee bewerkte kanalen opleveren: een blauw/geel kanaal en een helderheidskanaal. Hiervoor moesten natuurlijk ook de onderscheidende neurale mechanismen ontwikkeld worden, niet alleen de fotoreceptoren! De biologische voordelen van een dichromatisch boven een monochromatisch systeem maakten de evolutionaire druk kennelijk groot genoeg om deze ontwikkeling krachtig te bevorderen. Men schat wel dat een monochromatisch systeem maximaal ongeveer 100 stappen in helderheid kan coderen, en dat een dichromatisch systeem het aantal combinaties van helderheden en kleuren verhoogt tot ongeveer 10.000.
De grote meerderheid van de zoogdieren, inclusief huisdieren als hond en kat, is nog steeds dichromaat, volgens het bovengenoemde schema. Deze dieren zijn dus niet kleurenblind, maar hun kleuronderscheidingsvermogen is veel beperkter dan het onze. Onder de zoogdieren hebben alleen de mens en de apen van de ‘oude wereld’ (Europa, Azië en Afrika; bijv. de makaken) trichromatie ontwikkeld. Deze is tot stand gekomen door differentiatie van het l-gen in een m- en een l-gen. Men schat dat deze rond 50 miljoen jaar geleden is ontstaan, dus veel recenter dan de s- l divergentie, samenvallend met de afsplitsing tussen apen van de oude en nieuwe wereld (door het uiteendrijven van de Afrikaanse en Zuid Amerikaanse continenten). De opsines van m en l kegels verschillen slechts in een 15-tal aminozuren, waarvan slechts 7 bepalend zijn voor de verschillen in de absorptiespectra, die dicht bij elkaar liggen (Fig. 3); het grootste deel van dit verschil wordt zelfs bepaald door slechts 2 aminozuren. De toevoeging van het m-fotopigment (en de neurale circuits voor het groen/rood kanaal) betekent ten opzichte van de dichromaten wederom een enorme uitbreiding van de te onderscheiden kleur/helderheid combinaties, tot ongeveer 1 miljoen. (Moderne kleurmonitoren en digitale camera's kunnen miljoenen kleur/helderheid combinaties coderen, dus
| |
| |
in principe ruim voldoende voor ons visuele systeem!).
Met deze recente ontwikkeling tot trichromaten zijn mens en oude wereld apen allerminst kampioenen kleurenzien in het dierenrijk. Het is de laatste jaren duidelijk geworden dat zeer veel bij dag actieve niet-zoogdier vertebraten, zoals vissen, vogels en reptielen, vier kegelpigmenten hebben dus tetrachromaat zijn. Het vierde pigment is gevoelig voor ultraviolet (uv) licht, met een golflengte kleiner dan 400 nm. uv vormt een belangrijk bestanddeel van het invallende zonlicht, maar wordt door objecten meest geabsorbeerd en niet weerkaatst, uv-gevoelige kegeltjes zullen dus waarschijnlijk niet zozeer een rol spelen in de discriminatie tussen objecten, maar in het waarnemen van de aanwezigheid van de hemel en grote open ruimtes.
Volgens sommige (speculatieve) theorieën zijn de zoogdieren aanvankelijk tot dichromaten verworden omdat ze ontstonden tijdens de heerschappij van de dinosauriërs (dagdieren); ze moesten zich daarom aanvankelijk handhaven als nachtdieren, en pasten hun visuele systeem aan zeer lage lichtniveaus aan, waarbij mogelijk twee van de vier oorspronkelijke fotopigment-genen geïnactiveerd werden.
Vele invertebraten, zoals insecten, beschikken ook over vormen van kleurenzien. De meeste beschikken over uv, s en m fotopigmenten. De kennis hierover is echter nog vrij fragmentarisch.
| |
Defecten in kleurenzien
Zoals voor de hand ligt kan het kleurenzien, evenals de andere kwaliteiten van het zien, verstoord worden door ziekteprocessen die de ogen, en in het bijzonder de retina, aantasten. Op zulke verworven defecten, die allerlei aard kunnen hebben, wordt hier verder niet ingegaan. Ook het zeer zeldzame selectieve verlies van kleurgewaarwording door hersenbeschadigingen bespreek ik niet; de weinige rapporten hierover geven veel aanleiding tot controverse en zijn vaak nogal anekdotisch.
In het kader van deze inleiding zijn echter wel de aangeboren, genetisch bepaalde afwijkingen van het kleurenzien (‘kleurenblindheid’) uiterst belangwekkend. We zagen al dat de selectieve absorptiespectra van de 3 kegeltypen bepaald worden door het opsine (eiwit) deel van hun fotopigment. Voor het coderen van elk van de 3 opsines is een gen aanwezig in het erfelijke materiaal (het dna). Deze genen hebben vermoedelijk ook geen andere functies dan het maken van de fotopigmenten, en we hebben hier dan ook een uitgesproken geval van het ontstaan van een scherp omschreven functioneel defect door disfunctie van één gen.
Het gen voor het opsine van de (‘blauwgevoelige’) s-kegeltjes ligt op chromosoom 7. Defecten hiervan leiden tot ‘tritanopie’, maar deze afwijking is zeer zeldzaam (misschien 1 op 65.000 mensen). Het visuele effect
| |
| |
schijnt vooral een verwarring tussen blauwe en groene kleuren te zijn, naast een verlaagde gevoeligheid voor korte golflengtes.
Zeer veel vaker komen afwijkingen voor in de rood-groen discriminatie. De twee genen voor het l en m opsine liggen naast elkaar in het x-chromosoom, waarvan vrouwen twee exemplaren hebben maar mannen slechts één. Het feit dat de twee genen direct naast elkaar liggen en erg weinig van elkaar verschillen maakt ze zeer kwetsbaar voor herschikkingsprocessen (recombinaties) waarbij een van de genen verloren kan gaan of hybride genen (tussenvormen) kunnen ontstaan. Bij mannen komen zulke fouten het vaakst tot uiting, omdat ze maar één x-chromosoom, dus één set l en m opsine genen hebben. Wel 8% van hen vertoont een matig of ernstig rood-groen defect. Als het l-gen uitgevallen is, worden geen l-kegels ontwikkeld; dit heet protanopie. Zo iemand is minder gevoelig voor lange golflengtes (rood lijkt donker) en kan in het groen tot rood gebied zeer moeilijk kleuren van elkaar onderscheiden. Ontbreekt het m-gen, dan worden geen m-kegels gevormd. Dit heet deuteranopie, dat bijna het zelfde discriminatieprobleem oplevert. Protanopen en deuteranopen zijn dichromaten; ze hebben maar twee typen kegels. Mildere vormen van rood-groen kleurenblindheid zijn de protanomalie en deuteranomalie. Dit betreft mensen die wel trichromaten zijn, maar waarbij een van de m en l genen zo veranderd is dat de spectrale gevoeligheid van de twee gevormde fotopigmenten heel dicht bij elkaar ligt. Bij protanomalie zijn er geen normale l-kegels, maar twee typen m-kegels met iets verschillende absorptiespectra. Bij deuteranomalie zijn er geen normale m-kegels, maar wel twee, iets verschillend absorberende,
l-kegels. Deze anomale trichromaten hebben dezelfde soort problemen met kleurenzien als de dichromaten, maar in lichtere mate.
De praktische nadelen van rood-groen defecten moeten niet gebagatelliseerd worden. Moeilijkheden doen zich voortdurend voor bij het interpreteren van kleur-gecodeerde internetpagina's, landkaarten, verkeerslichten, het opmerken van een lichte verkleuring van de huid, of het kiezen van bij elkaar passende kleren. Ook de voor een normale trichromaat zo opvallende verschillen tussen kersen en bladeren of rijp en onrijp fruit (Fig. 1, kleurplaten) zijn voor een rood-groen defecte niet te zien!
| |
Zie jij wat ik zie?
Het bovenstaande leidt al snel tot de vraag wat voor kleuren een dichromaat dan eigenlijk ziet. Strikt genomen is dit soort vragen niet te beantwoorden, omdat niemand van een ander - normaal of abnormaal - kan weten wat die precies ziet. Toch zijn er wel aanwijzingen, die ondersteund worden door enkele (zeer zeldzame) gevallen waar het roodgroen defect is beperkt tot één oog, zodat de betreffende persoon de per- | |
| |
cepties via zijn afzonderlijke ogen kan vergelijken. Het lijkt er op dat deuteranopen in het golflengtegebied waar normalen rood en groen zien, in plaats daarvan een soort beige, geel en oranje zien.
Dit blijft natuurlijk een half antwoord op een feitelijk onbeantwoordbare vraag. De fysioloog kan het verband verklaren tussen kleuren en golflengtes, evenals de werking van de retina en, tot op zekere hoogte, de verwerking van golflengte/intensiteit informatie in de hersenen. Hij kan ook aantonen dat twee normale mensen de zelfde hoeveelheden rood en groen licht mengen om een gegeven testkleur ‘geel’ na te bootsen. De gewaarwording ‘geel’ als zodanig blijft echter een intrapsychische activiteit, die verder niet objectiveerbaar is of vergelijkbaar met die van iemand anders. Het blijft dus in theorie mogelijk dat deze voor de ene (niet kleurenblinde) mens niet gelijk is aan die van de ander. Dit is nauwelijks verder te onderzoeken, en vragen hierover zijn vrijwel zinloos, temeer daar benamingen voor kleurgewaarwordingen natuurlijk aangeleerd zijn, en daardoor genormaliseerd.
| |
Verdere oriëntatie
Zeer veel informatie over kleurenzien is te vinden op het internet, van populair tot specialistisch. Ik noem slechts enkele bronnen:
| www.mcw.edu/cellbio/colorvision/test1.htm |
| Geeft een fraaie indruk van de problemen van een deuteranoop (auteur: Neitz en collega's). |
|
| www.mcw.edu/cellbio/colorvision/colorvision.pdf |
| Een leuk overzichtsartikel uit de zelfde groep. |
|
| www.handprint.com/hp/wcl/wcolor.htm |
| Vooral bedoeld voor beeldende kunstenaars, vele hoofdstukken. |
|
|
