
Bloed
(1968)–Leo Vroman–
[pagina 105]
| |
IV Rode cellenStel je een micel van fosfolipide voor. In je plasma. Daar drijft het rond, een zacht kluitje moleculen met al die vette staarten binnenin, en al die waterlievende kopjes buitenop, naar het plasmawater kijkend. Blaas de micel nu eens op met plasma, om er een ballonnetje van te maken. Nu heb je een nieuw oppervlak gemaakt: inwendig, en doordat de inhoud van het ballonnetje nat plasma is, moeten ook langs het inwendige oppervlak de fosfolipide-moleculen hun kopjes uitsteken en niet meer hun staarten. Een soort van dubbellaag moet ontstaan, en de plasmaproteïnen, met hun hydrofiele en hydrofobe groepen, kunnen aan al die rangschikkingen deelnemen, zodat het hele ding eruit kan gaan zien als in figuur 38 op min of meer dwarse doorsnede te zien is. Daar heb je net zo ongeveer alles wat we van het rode-celmembraan afweten. Natuurlijk wordt het niet op zo'n primitieve manier gevormd. Trouwens, het heeft misschien wel een dubbele dubbellaag van fosfolipiden. Bovendien kan het niet erg egaal verdeeld zijn, want dan zou een rode cel bolvormig zijn en zouden de vette staarten een ononderbroken laag vormen die geen water zou doorlaten. Toch, zelfs uit de fijnste details van membraanstructuur die tot nu toe bekend zijn kunnen we nog niet verklaren hoe de cel zijn gekke vorm behoudt, water filtreert, natrium uit zich wegpompt of weghoudt en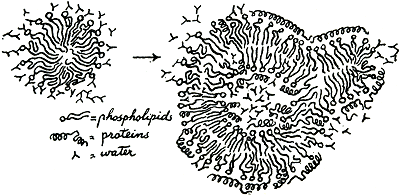
Figuur 38.
| |
[pagina 106]
| |
kalium binnenhoudt, zorgt dat hij niet aan andere cellen vastplakt en toch zichzelf zo zacht houdt, dat hij week als vloeistof door je capillairen stroomt. Microscopisten, chemici, immunologen en reologen kunnen daarvoor wel modellen bedenken waarmee ze zichzelf bevredigen maar niet elkaar. Ik beschouw nu mijzelf maar als een verbijsterde toekijker, maar al veel jaren voordat ik zelfs maar een eiwitmolecuul probeerde te naderen was ik al op een millimeter afstand van enige rode cellen geweest: als student, in Nederland, via een microscoop. Graag hield ik toen het gewebde koude voetje van een kikker vast, om door de microscoop naar zijn bloedvaten te kijken, de rode cellen langs te zien rennen in een kleine arterie, en om te proberen een enkele cel te volgen, hoe die bij een splitsing aangeland even aarzelde, een arteriole uitkoos, een langzame rit, een kronkelige capillair door die hem uitkneep van een schijf tot een cilinder, een kegel, een gek puntmutserig ding, en dan... opluchting! voort een wijder bloedvat in met ruimte om weer schijverig te worden, door de venules een vene in met levensgevaarlijke haast. Een kikker, zoals de meeste amfibieën, heeft grote ovale cellen met kernen. De onze zijn maar 7,8 micron in doorsnede (figuur 39). 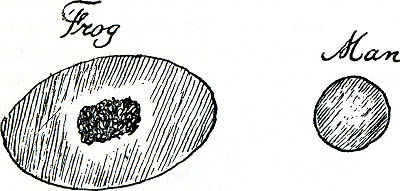
Figuur 39.
Dat is niet erg groot, maar kort nadat ik in New Brunswick begon te werken leken ze al zo groot als een kwartje, van al het kijken. Later, toen ik in moleculaire structuur begon te denken, groeiden de rode cellen opnieuw en ze zijn nu ongeveer drie meter hoog en hoorbaar, hoewel nog even geheimzinnig als ooit. De elektronenmicroscoop helpt zowel de cel als zijn geheimen te vergroten. In plaats van lenzen gebruikt hij hoepels van elektriciteit waardoorheen elektronen vliegen in plaats van licht. Het veld dwingt de elektronen door een | |
[pagina 107]
| |
brandpunt te gaan en door de meest transparante delen van je preparaat, dat zo een soort van geweldige schaduw werpt op een scherm. Ongelukkig genoeg mogen die elektronen natuurlijk nooit van hun rechte pad worden afgeleid, dus de ruimte rondom je preparaat is streng verboden toegang voor lucht en andere ronddolende moleculen.
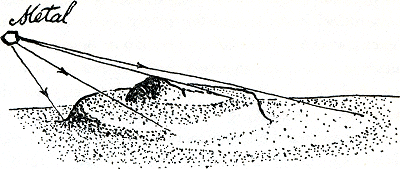
Figuur 40.
In het vacuüm dat dan ook nodig is, moet je preparaat worden bekeken onder omstandigheden zo gezond als blootshoofds op de maan. Al het water is weggekookt en alleen de mummie van het eens zo sappig dun plakje is over. Zelfs dan is het vaak nog veel te ijl om een duidelijke schaduw te vormen. Er bestaat een mooie manier om het meer zichtbaar te maken. De gemummificeerde plak, of zelfs een afgietsel daarvan, wordt in een luchtledige ruimte gelegd, en laag aan de horizon daarvan wordt een klein stukje metaal gehangen als een nietig zonnetje. Dat wordt heet gemaakt zodat het metaalatomen uitstraalt in alle richtingen, ook laag en schuin tegen de mummie; vooral de ‘zonnekant’ van de dikste plekjes zal veel metaal opvangen, en die zelfde, uitstekende plekjes zullen lange schaduwen werpen waar ze het achterland beschermen tegen de metaalstralen. Als dit schuine bestuiven klaar is, wordt het ding in de elektronenmicroscoop gelegd, waar de tegenstelling tussen de voor elektronen ondoorzichtige metaallagen en doorzichtige ‘schaduwen’, vooral als negatieve afdruk gezien, een verbazend realistisch beeld vormt. Een dorre woestijn bij avond in vogelvlucht gezien, met in de ondergaande zon de verwrongen dingen door hun water achtergelaten. Hoe afschuwelijk alles verminkt is door de vreselijke droogte en het blikke licht-niemand kan dat nog zeggen. Het verdorde membraan van een rode cel zal op deze manier vol dikke schijfjes zijn, verschillend lijkend voor verschillende diersoorten. Waren dat eilandjes van platte micel- | |
[pagina 108]
| |
len toen ze nog leefden? Misschien; dan is het ook in te denken hoe vetachtige stoffen zo goed in levende rode cellen kunnen binnendringen: door de hydrofobe centra van die micellen. Het zou wel leuk geweest zijn als we op zulke foto's ook poriën in zo'n membraan hadden kunnen zien, of kanaaltjes, die groot genoeg zouden moeten zijn om water door te laten maar niet bepaalde ionen of vooral moleculen. Onzin: zo simpel is de samenhang tussen vorm en functie nooit; dat zou even dwaas zijn als te verwachten dat de beweging van mensen hun huizen in en uit moet afhangen van hoe groot hun deuren zijn. We moeten uitvinden wat hun om te beginnen tot een bepaalde deur heeft geleid. De rangschikking van atomen langs de kromme ruggegraat van ieder molecuul membraaneiwit, en hun verband met de atomen in fosfolipidemoleculen en met de haren, kwasten, webben en knuppeltjes puur water die zich elke miljoenste seconde tienduizend keer vormen en weer vernietigen-dat zouden we allemaal moeten zien voordat we kunnen begrijpen waarom een natrium-ion heel andere avonturen over zijn tocht door een membraan heeft te vertellen dan een kalium-ion. | |
Functies van membranenSommige geleerden hebben het gevoel dat het de structuur van het membraan zelf is, die de structuur van het water dwingt, bepaalde stoffen in bepaalde richting te transporteren. Een rij watermoleculen, bijvoorbeeld, bij elkaar gehouden bijna zoals een magneet een hele keten spelden aan zich kan laten bungelen, zou in staat zijn een elektron door te geven van het ene naar het andere molecuul. Of tenminste, een molecuul zou zijn waterstofband verplaatsen en daardoor zou de volgende het ook doen, enzovoort, zoals een handdruk kan worden doorgegeven zonder dat ook maar een enkele hand het bijbehorend lichaam hoeft te verlaten. Een argument voor zo'n nogal passief transportsysteem is dat dode draden wol even goed het verschil weten tussen natrium- en kalium-ionen (wat toch heel moeilijk moet zijn) als levende (of misschien wel niet levende) membranen. Bovendien, zeggen die geleerden, als je aanneemt dat er werkelijk energie gebruikt wordt door het membraan voor transport, dan blijkt uit berekeningen dat voor zoveel energie in het membraan geen plaats is, en dat die zelfs niet in de hele cel aanwezig is. De meeste andere ge- | |
[pagina 109]
| |
leerden kijken liever naar de tegenovergestelde bewijzen, dat er aan selectief transport gewerkt moet worden en dat het membraan moet blijven eten om in leven te blijven. Ze geloven dat er een actieve natriumpomp is, die het natrium de cel uitpompt als er te veel binnenkomt. Als je een rode cel uithongert door hem in een oplossing te stoppen zonder glucose, beginnen al gauw de verkeerde ionen de cel in en uit te lekken. Wat dat allemaal bewijst, of wie gelijk heeft, dat weet ik niet; wat mij betreft kan de waarheid wel in het midden liggen. Waarom bijvoorbeeld niet geloven dat er energie nodig is voor het handhaven van een overigens ‘dood’ membraan? In de wereld van moleculen wordt het wel eens wat moeilijk, of zelfs belachelijk, een principieel verschil te zoeken tussen het levende ding, en het dode dat daaruit is ontstaan. Zelfs al is het dood, dan is het membraan toch beslist niet iets gewoons. Doe eens een druppeltje vers bloed op een voorwerpglaasje, en een druppeltje water ernaast, en bedek ze samen met een dekglaasje. Kijk dan door de microscoop waar de twee druppels elkaar raken en vermengd worden. Daar kan je de rode cellen zien veranderen: hun holle flanken beginnen te zwellen van het water dat de cellen opzwelgen, een beetje hemoglobine gaat al door hun opengesperde poriën naar buiten, en tenslotte zweven hun lichamen rond als kogelronde en toch lege, bleke blazen. Had je op tijd wat zout bij het water gedaan, dan hadden de cellen al gauw hun oorspronkelijke ingezogen schijfvorm herwonnen door hun teveel aan water weer te verliezen, en als je er teveel zout bij had gedaan, zouden de cellen zoveel water uit hun lichamen persen dat ze er kleine gekreukelde propjes van zouden worden, bezet met idiote stekels (figuur 41).
Figuur 41.
Ook deze walgelijke gebeurtenis kan je weer helpen door de te zoute oplossing met water te verdunnen: dan drinken de cellen zich terug tot de oude vorm. Doe er weer zout bij en de cellen zullen weer precies zo ineenkrimpen met dezelfde stekels op dezelfde plekken zelfs. De bedoeling van dit wrede spelletje is alleen maar dat we er conclu- | |
[pagina 110]
| |
sies uit trekken. Welke? Misschien, dat er allerlei onzichtbare structuren zijn niet alleen in het membraan maar binnen in de rode cel, die hem bij het krimpen specifiek en zichtbaar verwringen. Maar het is natuurlijk ook interessant dat zo'n rode cel, en elke andere cel, water opzuigt uit een omgeving die ‘wateriger’ is dan zijn binnenste, en water zweet als hij van binnen wateriger is dan zijn omgeving. Een antieke maar misschien wel handige manier om daarover te denken is: vergeet dat het water bestaat, en verbeeld je dat de daarin opgeloste stoffen rondzweven als de moleculen van een gas, en druk uitoefenen als een gas tegen het membraan. Die druk heet osmotische druk; je kan hem ook toniciteit noemen. Een oplossing van 0,85% keukenzout heeft ongeveer de osmotische druk die in onze cellen voorkomt en wordt daarom isotonisch genoemd. Daarin kan je de cellen dus een heel tijdje laten rondzweven zonder dat ze opzwellen of ineenfrommelen. Als je de vorige paragraaf in een wetenschappelijk taaltje wil uitdrukken, kan je zeggen dat rode cellen hemolyse vertonen in hypotonische, en crenering in hypertonische oplossingen. Volgens een oude theorie is de toniciteit en zelfs in detail de samenstelling van zouten in ons plasma sprekend die van zeewater, of sterker nog, ons plasma zou nog het voortgeërfde zeewater zelf zijn. Slijmerig kropen we lang geleden uit de oceaan het land op, waar onze huid, zich verhardend tegen de zon, de droge lucht en de zouteloze regen, zich stevig sloot rondom het bitter zoute zeewater dat we binnen onze lijven uit de zee hadden meegesleurd. En nog altijd lopen we onze cellen te weken in dit namaaksel van een of andere voorhistorische Indische Oceaan. Dat is blijkbaar nog altijd goed genoeg. Wat hebben onze rode cellen daar dan op aan te merken? Want ze staan erop het kalium daaruit te verzamelen en het natrium terug te sturen naar de zee rondom ze. Is de scheve verhouding tussen hun elementen misschien nodig voor rode cellen om te kunnen werken? Niks hoor: hond en kat hebben juist een laag kalium- en hoog natriumgehalte in hun rode cellen, terwijl rat en konijn daarin zijn zoals wij. Het kaliumpeil van sommige schapen is hoog, van andere laag. Het is dus moeilijk, in het ingewikkeld gescharrel van het membraan met ionen iets van belang te zien. Wel schijnt er een samenhang te bestaan tussen de fosfolipide-compositie van de rode-celmembranen en de kalium: natriumverhouding van de celinhouden. Er wordt | |
[pagina 111]
| |
dan ook tegenwoordig veel in bekers vol verschillende zoutoplossingen geroerd die door lagen vet en fosfolipiden van elkaar gescheiden zijn. Alweer schijnt het membraan een dode uitdrukking van het leven. | |
BloedgroepenDe onzichtbare structuur van het celmembraan is ook tot uitdrukking gekomen in vreemde ontdekkingen van bloedgroepen. Blijkbaar heb je een eeuwige drang geërfd om bij elke rode cel wat vettige en suikerige moleculen te maken, die waarschijnlijk met hydrofobe banden op de vette plekken van het rode-celmembraan vastplakken. Zo kan je behoren tot, bijvoorbeeld, groep o, a, b of ab, afhankelijk van de twee genen (een per ouder) die je georven hebt. a+a of a+o maken je tot groep a, b+b of b+o maken je b, a+b ab, en alleen van o+o word je o. Tegen de stof die je zelf niet maakt, vormt je trouwe maar stomme lichaam een antilichaam (een eiwit) dat je moet ‘beschermen’ tegen die vreemde maar volkomen onschadelijke stof. Volkomen goedgezinde rode cellen van een groep b-heer, getransfuseerd in een groep a-man, zullen onmiddellijk worden bekleed met het eiwit anti-b, zodat ze verstikt aan elkaar zullen plakken en hemolyseren; hun dode kleine lijven zullen de capillairen van 's mans nieren verstoppen tot het hele lichaam van a mee moet sterven, typisch het slachtoffer van een menselijk zinloze oorlog. Het abo-groep-systeem en nog enige andere zijn al zo goed bekend en zo makkelijk na te gaan dat ze tegenwoordig niet veel dergelijke ongelukken meer veroorzaken. Maar nog altijd worden er ‘nieuwe’ gevonden. Zoals de stollingsfactoren, worden ook de meest algemeen voorkomende bloedgroepen het laatst ontdekt. Want neem eens aan dat mevrouw Xippepaai, toen ze nog maar een eitje was, door een kosmische straal in een gen werd geraakt waar voordien nog nooit iemand geraakt was, en waar de eigenschap had gezeten die verantwoordelijk had moeten worden voor het maken van een bepaalde stof X (genoemd naar mevrouw Xippepaai, de enige die die stof juist niet maken kan). Mevrouw X is opgegroeid en getrouwd. Ze wordt zwanger. Het gaat best met dat embryo; door de vader gezegend met normale genen, begint het zijn eigen rode cellen te maken met X-stof erop (sorry hoor, die letter heeft niets met het X-chromosoom | |
[pagina 112]
| |
te maken!). Kort voor zijn geboorte lekken een paar van die cellen of stukjes ervan door de placenta heen en komen in de bloedstroom van de moeder terecht. Terwijl die, bij wijze van spreken, aan de ene kant een normale baby geboren laat worden, maakt ze aan de andere kant antilichamen tegen de stof X. Een paar dagen later besluit het hospitaal om de gewone onbegrijpelijke reden, haar op te vrolijken met een transfusie. Haar bloedgroep wordt bepaald: haar rode cellen klonteren in serum van B-mensen, maar niet merkbaar in serum van A-mensen. In een beetje bloed uit een keurige fles vol A, veroorzaakt een druppel van haar serum maar nauwelijks een beetje geschift. Toevallig is het juist maandagochtend, en de knaap die deze bepalingen doen moet heeft een barstende hoofdpijn waardoor de hele wereld geschift lijkt. Hij vervloekt zijn ogen, zegent de fles bloed en de transfusie wordt begonnen. Al gauw krijgt mevrouw X rillingen: transfusiereactie! Gelukkig heeft ze verder geen bloed nodig, want het beetje dat ze al ontvangen had werkt net als een injectie dode bacteriën of zo die we zo nu en dan moeten hebben om onze produktie van antilichamen te versterken. Mevrouw X blijft dan ook nog een hele tijd ijverig anti-X maken, en haar serum of plasma zou nu duidelijk alle soorten rode cellen van de wereld aan het klonteren brengen behalve die van haarzelf. Nou ja, ze wordt beter, en twee jaar later heeft ze weer een kind. Ook dat lijkt eerst gezond, maar laat in de middag begint zijn huid wat gelig te worden, en het wit van zijn ogen, en het lijkt niet zo actief meer. Diezelfde laboratoriumknaap, sindsdien van de drank af, komt kijken en neemt wat bloed van het kindje. Een druppel doet hij op een voorwerpglaasje en strijkt hem uit met een ander glaasje, zo (figuur 42): 
Figuur 42.
| |
[pagina 113]
| |
De rest van het bloed doet hij in een reageerbuisje. Teruggekomen in zijn lab, legt hij het uitstrijkpreparaatje dwars op twee ondersteunende staven boven een gootsteen en giet er een donkerpaarse vloeistof op: Wright's kleurstof, een mengsel van eosine (een rode zurige stof), methyleenblauw (een meer basische, blauwe stof) en methylalcohol. De alcohol breekt allerlei waterstof- en hydrofobe banden in de eiwitten van de cellen en het plasma waarop ze liggen uitgespreid, zodat ze vast worden en onoplosbaar in de druppels water die daarna worden toegevoegd. Nu hecht het eosine zich vooral vast aan het meer basische protoplasma van de cellichamen, en het methyleenblauw gaat vooral op de nucleïnezuren van de kernen zitten. Dan wordt het glaasje afgespoeld met water en onder de microscoop bekeken. De witte cellen zien er nu zacht roze of bleek paarsig uit met donker bruinblauwviolette kernen, de rode cellen oranjeroze, en tussen alles door ligt een fijn sproeisel van paarsgestippelde bloedplaatjes. Maar overal in het uitgestreken bloed van deze baby vindt de goed voorbereide laboratoriumwerker cellen die een heel speciale betekenis hebben, ronde cellen met grote ronde kern en grijs protoplasma die hij pronormoblasten noemt; als ik je nu ook nog vertel dat een normale rode cel vaak normocyt wordt genoemd, en dat ‘blast’ komt van een Grieks woord dat ‘spruiten’ betekent, dan kan je wel raden wat de naam van die cel betekent: een heel jonge overigens normale rode cel. Zijn juiste plaats is niet in het bloed maar in het beenmerg. Dan is er ook nog een soort cel die meer op een gewone rode cel lijkt, het ronde lijf al oranje-achtig en blijkbaar betrapt bij het maken van zijn eerste hemoglobine, maar nog steeds met een nu bijna zwart gekleurd, rond dingetje erin: de krimpende kern. Deze cel is een normoblast; het kleine zwarte bolletje was al klaar om de cel uit te glippen. In het reageerbuisje intussen is het bloed gestold, het stolsel is samengetrokken en een geel, veel te geel serum eromheen wordt zichtbaar. Wat is er allemaal met dit kindje gebeurd? Zoals het eerste, heeft ook hij van zijn vader het gen gekregen om X-stof op zijn rode cellen te maken, maar zelfs vóór hij een bevrucht eicelletje was, had dat oudere broertje zijn moeder met wat X-stof dragende rode cellen ingespoten-zoals je weet. En toen had die moeder ook nog die hele transfusie van normale, dus X-dragende cellen gekregen of misschien zelfs gekocht, zodat ze flink immuun was geworden tegen X. Toen | |
[pagina 114]
| |
deze tweede baby zijn eigen rode cellen begon te maken begon de moeder daar meteen anti-X tegen te fabriceren in een woest oplaaiende oorlog tegen haar eigen inwendige kind, wiens nieuwe rode cellen met haar anti-X werden bekogeld, beplakt en verstikt zodra ze gevormd werden. Langzamerhand is het baby-beenmerg wanhopig geworden,
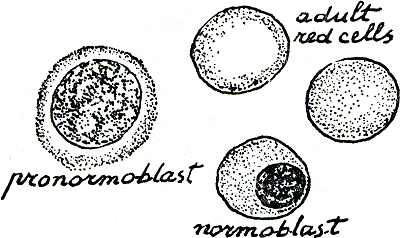
Figuur 43.
en in plaats van rustig volgroeide rode cellen het bloed in te sturen, probeert dat het nijpend tekort in te halen door steeds jongere cellen uit te storten. De lever, die tegen deze tijd al bereid had moeten zijn om de rode-cellenproduktie maar aan het beenmerg over te laten, is nog steeds op volle gang, en zelfs de milt, die al een half jaar geleden had moeten ophouden, gooit nog halfgare rode cellen het bloed in. Uit de door anti-X aangevallen en gewonde cellen barst intussen het hemoglobine dat nu moet worden omgezetin galpigmenten: bilirubine bijvoorbeeld, dat door de lever moet worden opgenomen om gal van te maken. Maar de onhandelbaar zware lading geel goedje maakt van het plasma waar het niet uitkan een bijna oranje vloeistof. Nu zijn de doktoren werkelijk bezorgd, en laten de labknaap telkens wat bloed van de baby nemen om het bilirubinepeil af te roepen. Een specialist in de bloedbank staat al klaar: als de bilirubine boven een bepaalde concentratie stijgt, moet hij al het bloed van dit kind door een even grote hoeveelheid onschadelijk bloed vervangen-als hij dat ooit kan vinden. Laten we aan dit verhaal maar een gelukkig en helemaal niet onmogelijk einde maken: het bilirubinepeil stijgt niet meer, begint dan te dalen, steeds minder kerndragende rode cellen zijn in de uitstrijkpreparaatjes te vinden, de baby wordt levendiger, en juist als moeder en kind klaar staan om naar huis te gaan, komt me daar een geleerde | |
[pagina 115]
| |
uit de grote stad binnen en biedt het nu geheel straatarme vrouwtje duizend gulden voor een flesje van haar bloed, om daar spreeuwecellen mee te laten klonteren. Meestal is het noch zo'n zeldzame groep, noch het a-b-o-systeem dat narigheid veroorzaakt, maar het Rh-systeem. Rh betekent Resusaap. Om alles maar niet zo ingewikkeld te maken als het is, zal ik alleen maar zeggen dat je Rh-positief bent als jij en de Resusapen eenzelfde stof op jullie rode cellen maken. Je kan dat makkelijk genoeg uitvinden: spuit een of ander beest in met Resusaperodecellen. Dan maakt dat beest daar antistoffen tegen en zal zijn serum niet alleen de cellen van Resusapen laten klonteren maar ook die van Rh-positieve mensen. Schaam je vooral niet om alleen maar zo'n kleinigheid als een bloedgroepstof met wat apen gemeen te hebben. De gekste dieren hebben a-stof. Sommige planten hebben zelfs zoiets als a, en andere iets dat verdacht veel op anti-a lijkt. Mensen van Scandinavië en Australië hebben meestal a, Aziaten b. Wat moet dat nu wel betekenen? Ik heb er geen idee van. Wel zijn die bloedgroepmoleculen allemaal nogal klein. Best mogelijk dat ze uit een handjevol suikers, vetten, vaste voorwerpen en enzymen sneller en makkelijker gevormd worden dan andere moleculen. Als bacteriën dat vroeger al deden, en planten daar dan echte antistoffen tegen gevormd zouden hebben net als wij, dan is de diepe betekenis van de merkwaardige gelijkenissen misschien alleen maar dat planten en dieren aan gemeenschappelijke wetten gehoorzamen. Mens en plant, hand in hand. Nou dan, tot zover het oppervlak van de rode cel, die gladde maar chemisch bonte buidel van bijna vloeibare vervormbaarheid en altijd weerkerende rare vorm, volgeplakt met misschien voor geen twee mensen dezelfde kleine zoete moleculen, die ionen herkent en eiwitten zo omhult dat ze binnen de buidel zestig maal langer kunnen leven dan in het plasma daarbuiten. En zo komen we dus in de rode cel terecht. | |
Waar komen ze vandaan?Juist was ik van plan een gezamenlijk tochtje door de rode cel te organiseren, toen ik bedacht dat ik je daarbinnen alleen maar een stel tegenstrijdige leugens zou kunnen laten zien. Misschien kan ik je be- | |
[pagina 116]
| |
ter vertellen hoe de rode cel zo geworden is, ook al weet niemand hoe hij is. Achteruit lopend, de jeugd van een rode cel in, door zijn geboorte heen het leven van zijn ouders in en zijn voorouders, verder terug waar ook het hele mensenlichaam eromheen jonger en kleiner wordt, een cel, terug door de ouders van die eicel en de voor-voorouders van alle zoogdieren, vallen we eindelijk achterover de nevel in waaronder het ontstaan van de wereld verborgen schijnt te zijn-een nevel van het verleden die niets dichter is dan de nevel van nu waarin de geboorte van elke cel elke dag is gehuld. Je kan een volwassen rode cel leren herkennen (en hem zelfs een gematureerde erytrocyt leren noemen) met je microscoop in een minuut, en het verschil tussen een normoblast en enkele verwarrende witte cellen leren in ongeveer tien minuten, maar het zal je dagen kosten voordat je een jonge pronormoblast hebt leren onderscheiden van een jonge soort witte cel (een lymfoblast), en experts hebben een heel leven nodig gehad om te leren dat nog jongere cellen gewoon niet uit elkaar zijn te houden. Of microscopen zijn daar nu eenmaal niet goed genoeg voor, óf alle bloedcellen stammen af van een primitieve troep voorvaderen, hurkend in ons beenmerg en allemaal aan elkaar gelijk op het tijdelijk toeval na, dat ze de ene dag wakkerschrikt om een of andere witte cel te baren, een week later om een nest pronormoblasten te werpen. In dat geval is het toeval zelf het geheimzinnigst, want het is zeker niet blind: elk soort cel wordt gemaakt naargelang de behoefte. Dat zieke kind, en elk mens met gezond beenmerg, reageerde op het verlies van rode cellen door meer rode cellen te maken, geen witte. Waarschijnlijk wordt de vorming van erytrocyten (erytropoiese) aangespoord door een bepaalde stof (erytropoietine) waar ik het later nog graag even over wil hebben. Eerst iets over verschillende manieren om het leven van een rode cel te volgen. De pronormoblast is zo iets (figuur 44): 
Figuur 44.
| |
[pagina 117]
| |
Zoals ik al zei, weinig protoplasma nog, en pas een begin van hemoglobinevorming. Meer en meer wordt dan, via kern-dna en korreltjes protoplasma-rna, hemoglobine gevormd, tot met de kern ook dat hele proces doodloopt. Wat overgeschoten nucleïnezuur-eiwitcomplex, een spookachtig spinrag, blijft nog even en vormt nog wat hemoglobine. Dat web kan met kleurstoffen worden gekleurd en de cel wordt dan een reticulocyt genoemd. Als je wilt weten of iemand anemisch is geworden doordat hij bloed heeft verloren of doordat hij te weinig nieuw bloed maakt, kan je een druppel van zijn bloed met zo'n kleurstof mengen en dan uitstrijken op een glaasje, en dan tellen hoeveel van zijn rode cellen op de duizend er reticulocyten zijn. Veel meer dan tien, dus meer dan 1 percent? Dan heeft de patiënt waarschijnlijk een normaal reagerend beenmerg en bloedt inwendig. Er zijn nog ruwere en toch behulpzame metingen die je op hem kunt doen. Neem wat van zijn bloed, doe er wat calcium-onttrekkend poeder (oxalaat bijvoorbeeld) of een druppeltje heparine (een lang molecuul, uit lever gemaakt) bij om stolling te voorkomen, giet het in een liefst nogal nauw stevig buisje, en centrifugeer het flink hard. Kijk daarna met een liniaaltje hoe hoog de kolom samengepakte rode cellen is en hoe hoog de kolom plasma die daar boven staat. Bij normale mensen moet de verhouding tussen die twee ongeveer 2:3 zijn; je kan dus zeggen dat onze rode cellen ongeveer veertig percent van al het bloed in beslag nemen, of, wetenschappelijker, dat de hematocriet ongeveer veertig percent moet zijn. Giet ook een afgemeten hoeveelheid verdund azijnzuur bij een afgemeten hoeveelheid bloed van de patiënt. Daarvan hemolyseren de rode cellen en wordt het vrijkomende hemoglobine een bruine smeerboel, waarvan je de bruinte kan vergelijken met een gekochte bruintestandaard of met een duurder doorlichtend apparaatje meten en omzetten in sterktewaarde van het oorspronkelijke hemoglobine. Nu heb je dus twee getallen: hematocriet en hemoglobine-concentratie van het hele bloed. Daaruit kan je dus ... berekenen; goed zo.Ga naar voetnoot* Je kunt ook werkelijk tellen hoeveel rode cellen er zijn in een bepaald volume van 's mans bloed. In principe is dat heel makkelijk: Neem een ‘rode-celpipet’, dat is een glazen buisje met een gezwel dat | |
[pagina 118]
| |
een ongeveer honderd keer zo groot volume heeft als het steeltje, dat daarentegen een schaalverdelinkje rijk is, zo iets: (figuur 45) 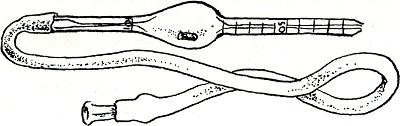
Figuur 45.
Maak een rubber slangetje vast aan het onverdeelde uiteinde zoals op het plaatje, en zuig bloed van de patiënt op tot het 0.5 streepje. Die handigheid leer je in drie vloekvolle dagen. Veeg de buitenkant af (weer een dag voorbij vol ellende) en zuig een soort onschadelijke verdunningsvloeistof op tot het 101 streepje zodat de hele pipet nu vol 200× verdund bloed is (daar gaat de rest van je week). Meng de inhoud een tijdje; het rode kraaltje in het pipettebuikje helpt daarbij. Laat nu enige druppels naar buiten lopen, veeg weer de buitenkant af, en laat de volgende druppel een ‘telkamer’ vullen. Dat is een duur stuk glas, zo gemaakt dat vloeistof een bepaalde dikte heeft als die zich tussen het stuk glas en een dekglas verspreidt over een in allerkleinste ruitjes verdeeld oppervlak. Laat de cellen rustig op de ruitjes neerzinken en tel dan hoeveel cellen je ziet in een bepaald aantal ruitjes. Nu weet je oppervlak en hoogte van het 200× verdunde bloed waarin je nu ook het aantal cellen weet, en dan maar uitrekenen. Als die man normaal bloed heeft moet je ongeveer komen op vijf en een half miljoen per kubieke millimeter van het oorspronkelijke bloed; als die man tenminste kalm was en jullie leven niet al te hoog boven de zeespiegel, anders meer. Maar zelfs een volmaakte analist heeft altijd tien percent kans dat zijn resultaat bijna een miljoen hoger of lager uitkomt dan het juiste getal-en niet alle analisten zijn volmaakt. Om zich aan die menselijkheid te ontworstelen kopen veel laboratoria tegenwoordig dure machines, voornamelijk nogal rechthoekige dozen die een monstertje bloed inslikken en dan uit een andere opening een gedrukt papiertje produceren, daarbij hun eigen fouten zowel als die van jou verbergend. Alleen door tijd te besparen geven ze je een groter aantal kansen om de waarheid te benaderen. En dan nog is | |
[pagina 119]
| |
de verleiding groot om het aardigste getal te kiezen. Stel dat je met een druppel bloed drie tellingen doet en de resultaten zijn: 4,0; 4,1 en 5,2 miljoen. Denk je dan niet meteen: die 5,2 zal wel een vergissing zijn, want die eerste is bijna hetzelfde als de tweede? Of, even verkeerd, ‘ik zal er nog eentje doen, als die dan toch dicht bij de 5,2 is neem ik maar het gemiddelde van alle vier’? Veel labs zijn helemaal niet streng met hun getallen of hebben zelfs geen tijd of zin voor meer dan een enkele waarneming. Veel onwetenschapppelijke medici geloven alleen in de juistheid van getallen die ze denken te begrijpen. Maar goed; aannemend dat je metingen goed waren, kan je uit het hemoglobine-gehalte van het hele bloed, uit de hematocriet en het aantal rode cellen per kubieke millimeter, het gemiddelde volume en de gemiddelde hoeveelheid hemoglobine van een rode cel berekenen. Te kleine of te grote, te volle of te lege cellen hangen samen met allerlei ziekten (en lange woorden), en bijvoorbeeld met de haast waarin de cellen gevormd worden. Maar er zijn elegantere manieren om uit te vinden wanneer in de patiënt erytrocyten worden geboren en gedood, waarvandaan ze komen en waarheen ze moeten gaan. Een paar van die manieren heb ik in het Mount Sinai Hospital geleerd, voor ik terugkroop in mijn eigen kleine stollingswereld. | |
Brandend etiketZodra ik Mount Sinai inkwam werd ik voorgesteld aan een lange, lage kelder vol kamers, de meeste door machines bevolkt die ik nog nooit had ontmoet. Helgele huiveringen stroomden over de zwarte gladde gezichten van stralingtellende machines omlaag; dinosauriërs zwaaiden langzaam hun gietijzeren koppen rond; ‘Danger! Radioactive!’ stond er op de deuren, de kasten, de flessen; en dan waren er ook nog analisten die de gewoonste dingen deden op de ongewoonste manieren, tafels dekten met boterhampapier, gewone machteloos lijkende flesjes aanpakten met handschoenen die daarna moesten weggegooid in een bepaalde prullenbak, ieder gebruikt pipetje of reageerbuisje dadelijk weglegden of in een bak waar ‘cold’ of in een waar ‘hot’ (radioactief) op stond. Zelfs de patiënten werden hier behandeld alsof ze meer dan de gebruikelijke bijrol speelden. Een grote kamer stond ze te wachten met de gordijnen omlaag, een of twee telmachines al traag tikkend, en twee tamme dinosauriërs aan hun dikke | |
[pagina 120]
| |
geïsoleerde slappe lijnen aan weerskanten van een onfatsoenlijk half opgemaakt bed neerstarend op de witte lakens. Naast het bed een klein wagentje waarop een onaantrekkelijke tentoonstelling van porties radioactieve ijzer-oplossing, een rek vol buizen en een stapel injectiespuiten was gereserveerd. 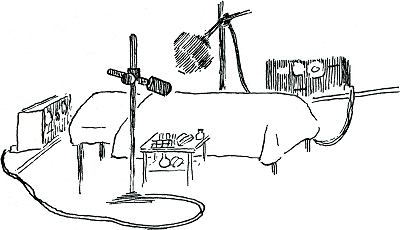
Figuur 46.
De hoofdpersoon van de eerste voorstelling die ik mocht meemaken bleek een klein vriendelijk oud vrouwtje te zijn in een kimono die niet in staat was het zwangerige van haar buik te verbergen. Ze kwam luisterend op met een dokter en een analiste tegen haar pratend: ‘Dus nou begrijp je alles toch wel hè liefie? Dat dit de hele dag zal duren?’ ‘Maar je hoeft niks te doen dan in bed liggen.’ ‘Ja, wij doen de rest wel.’ ‘Die machines gaan gewoon naar die grote gekke milt van je staan kijken; en naar je lever.’ ‘Om te zien wat die met je rooie cellen uitspoken. Want als je niet beslist moet, wat zou je me daar dan rondlopen met zo'n bakbeest in je buik, dat ze zeggen nounou en op die leeftijd nog, ja toch, dat is nergens voor nodig.’ Dat was eigenlijk wel waar: een milt die heel groot is en zelfs erytrocyten maakt kan meestal worden weggehaald zonder een gevaarlijke daling in het aantal rode cellen te veroorzaken. Maar in ieder geval was het belangrijk, ontstaan en vergaan van de rode cellen in zo'n patiënt te volgen. Daarom gaven we het brave bebbetje een gemeten hoeveelheid radioactief ijzer (Fe59); niet als een gewone oplossing, want een vrij | |
[pagina 121]
| |
ijzer-ion kruipt gewoon door allerlei soorten oppervlakken heen, ook door die van de bloedvatwanden, en we zouden het helemaal uit ons oog verliezen tenzij het op tijd zou worden gegrepen door een heel bepaald, ijzergrijpend eiwit, transferrine geheten. Daarop vastgezeten zou het ijzer kalm meekomen met de bloedstroom. Voordat het werd ingespoten werd daarom het ijzerzout eerst een tijdje gemengd met een oplossing van gezuiverd, steriel transferrine. De dame was intussen naar bed gegaan, waar het ene monster zijn straalgevoelige hoofd tegen haar milt hield gedrukt, terwijl het andere ondier ernstig aan haar lever probeerde te snuffelen. De twee telmachines waarmee elk dier verbonden was, waren al lang bezig langzaam de ‘achtergrond’ te tellen: de zwakke radioactiviteit van een lichtgevend polshorloge hier, een atoombom daar, en van kosmische stralen overal die ons allen eeuwig en pijnloos vanuit de eeuwigheid doorboren. Nu werd er wat bloed uit de linkerarm van de dame genomen; de naald moest in de vene blijven steken voor later en werd gekurkt en met plakverband tegen haar huid vastgezet. In een vene van haar rechterarm werd nu het radioactief ijzer-plus-transferrine ingespoten, daarna weer bloed genomen van haar linkerarm, niet alleen dadelijk maar ook met steeds langere tussenpozen na de ijzerinspuiting. Het bloed werd in buisjes gedaan waarop 30″, 1′, 2′, 4′, enzovoort werd geschreven. Die werden allemaal gecentrifugeerd, en een kubieke centimeter plasma van elk werd in een soort loden putje neergelaten. Daarin, door lood beschut tegen tenminste het zachtste radioactieve kabaal daarbuiten, telde dan een gevoelige bodem de activiteit van het plasma daarbinnen. De eerste buisjes plasma waren flink ‘heet’: de machine, afgesteld om bijvoorbeeld in totaal 10240 stralen te tellen en dan te stoppen en te zeggen hoe lang hij daarover gedaan had, was in enkele minuten klaar met de plasma's van vlak na de injectie. Maar plasma van het bloed dat ongeveer twee uur later was genomen, maakte dat de machine maar nauwelijks sneller tikte dan zonder iets; ook de rode cellen waren ‘koud’, zoals aldoor al. Het monster dat aan de milt stond te snuffelen werd daarentegen heel opgewonden: zijn telmachine ratelde vlugger en vlugger om ons te vertellen dat veel van het radioactieve ijzer in de milt was terechtgekomen. In een andere kamer werd intussen een beetje bloed-van-vóór-haar-ijzerinjectie steriel met steriel natriumchromaat gemengd waarin het chromium-atoom radioactief was. De rode cellen slurpten toen | |
[pagina 122]
| |
onmiddellijk dat brandend etiket op. Na een tijdje werd het chromium verder voor ze ondrinkbaar gemaakt (door er vitamine C bij te gieten), en daarna werden de cellen een paar maal in telkens ververste zoutoplossing gecentrifugeerd, gewassen heet dat; toen werd hun overgebleven en dus min of meer inwendige radioactiviteit gemeten en werden ze teruggespoten in mevrouws rechterarm. Kort daarna werd weer eens een beetje bloed uit haar linkerarm genomen en werd de radioactiviteit ervan gemeten. Denk je nu in dat een assistente met alle resultaten van tot nu bij je komt, en dat er ook nog andere metingen bestaan waarvan ik je al eerder vertelde. Wat kan je daaruit dan wijs worden? Nou, je kunt uitrekenen hoeveel bloed de dame heeft. Want uit volume, hematocriet en radioactiviteit van het terug-ingespoten bloed weet je hoeveel volume rode cellen van hoe grote activiteit je hebt ingespoten. Ook hoor je de hematocriet van de dame zelf te meten, en je hebt daarnet ook de activiteit na die injectie gemeten, waaruit je kunt berekenen hoeveel de activiteit van de ingespoten rode cellen verdund is door in haar hele lichaam gemengd te raken met haar eigen onactieve cellen. Bijvoorbeeld, als je vijf kubieke centimeter rode cellen met activiteit a hebt ingespoten, en later haal je uit de gemengde dame vijf kubieke centimeter rode cellen (je hebt immers het plasmavolume weggerekend met die hematocriet) met activiteit van een honderdste a, dan moet haar hele bloedsomloop 100×5 kubieke centimeter rode cellen bevatten. Als haar hematocriet twintig percent is, moet ze honderd gedeeld door twintig, maal 500 ofwel twee-en-een-halve liter bloed bevatten. Dat is nog eens een heel wat nettere manier dan die van een paar honderd jaar geleden, toen al het bloed verwijderd moest worden om een vrij ruw idee te krijgen. Tegen de tijd dat al die buisjes bloed genomen waren was de dag bijna vergaan en onze dame uitgeput. De volgende dag, en dagen, en toen nog één maal per week, werd bloed van haar genomen en daar de radioactiviteit van bepaald. Maar de getallen betekenden nu iets ingewikkelds: het mensje bevatte immers twee soorten activiteit tegelijk-van het chromium en van dat ijzer daarvoor. Hoe die uit elkaar te houden? Tegenwoordig kan dat bijna automatisch, met een soort filter die de twee elementen vanwege hun verschillende soorten straling kan scheiden. Maar in mijn tijd, twaalf jaar geleden, deden we dat anders. Elk radioactief element heeft zijn eigen halveringstijd. | |
[pagina 123]
| |
Van een element dat een halveringstijd van 13,23 dag heeft, verliest elke willekeurige hoeveelheid in 13,23 dag de helft van zijn activiteit. Als je dus een mengsel van twee actieve elementen hebt is de snelheid waarmee dat zijn activiteit verliest afhankelijk van de verhouding tussen de twee elementen die elk hun eigen verzwakking bijdragen. Daarom konden we het best de activiteit van het damesbloed met het actieve chroom en ijzer twee maal meten, met een tussenpoos van een paar weken; onze wiskundige kon toen met behulp van een paar differentiaalvergelijkingen uitvissen hoeveel wat wanneer waarin. Zo vonden we, dat op de dag na de inspuitingen, onze dame nog niets van ons ijzer in haar bloedsomloop had toegelaten, maar twee dagen later verschenen de eerste rode cellen met een actief ijzeretiket, daarna snel meer, terwijl al sedert de eerste dag het chroom, en daarmee waarschijnlijk de cellen die het chroom-etiket hadden gedragen, minder werden. Binnen korte tijd bereikte de ijzertelling zijn hoogtepunt en begon toen ook langzaam te dalen. Wat heb je hieraan? En aan de voorafgaande alinea's? In ieder geval, als je voor dokter durft te spelen, kan je afleiden dat de milt rode cellen maakte, want die werd immers ‘heet’ van het ijzer en dat verscheen nog geen drie dagen later al in de rondstromende erytrocyten. Jouw beenmerg kan dat, tussen twee haakjes, even vlug. Dat het chroom zo kort na de injectie al begon te dalen kan betekenen dat het meteen gedeeltelijk van de ingespoten rode cellen afviel en ergens buiten het bloed verdween, of dat er meteen heel wat van de ingespoten cellen zelf stierven en ergens verdwenen. Dat zou ook geen wonder zijn, want we hebben zo maar wat van haar bloed, dat cellen van alle mogelijke leeftijd moet hebben bevat, bij dat chromiumzout in het flesje gedaan, en sommige van die erytrocyten waren dus waarschijnlijk al klaar om te overlijden toen we ze vingen; terug-ingespoten, deden ze dat dadelijk. Voor die ‘fout’ kan een correctie worden berekend; en natuurlijk ook voor het vanzelf zwakker worden van de ingespoten radioactieve elementen zelf. Nou dan, die geschiedenis van onze oude dame had een gelukkig einde (wat een idiote uitdrukking; ik bedoel alleen maar dat hij zich op een gelukkig ogenblik kan laten afbreken): min of meer dank zij onze resultaten kon de milt worden weggenomen en daar ging ze van vooruit. Ik geloof dat het overdreven orgaan meer rode cellen opvrat dan het maakte, en dat ze daardoor veel korter dan normaal bleven leven. | |
[pagina 124]
| |
Hemoglobine en de levensduur van een rode cel waar bet in zitAls je normaal bent, leven je erytrocyten ongeveer 120 dagen. Dat kan je ook zo zeggen: ‘elke vier maanden heb ik al mijn rode cellen verwisseld voor nieuwe’. Of (even uitrekenen; kijk het vooral na): elke seconde moet ik er twee miljoen maken, en vermoorden. Maar goed dat ons aantal witte cellen niet veel meer dan een duizendste is van het aantal rode cellen, want de witte leven waarschijnlijk niet veel langer dan een week. En elk vrij rondstromend eiwitmolecuul in ons bloed leeft maar enkele woeste dagen en nachten. En toch, als een erytrocyt eenmaal volwassen is en ophoudt hemoglobine te maken, leeft hij nog 115 dagen langer; de hemoglobine binnen in hem ook. Het moet daar een gezond leventje zijn voor eiwitten, in die cel. Daar zijn namelijk ook enige enzymen in, bijvoorbeeld die welke het celmembraan waarschijnlijk telkens moeten repareren. En er leeft er een, methemoglobine-reductase geheten, dat zuurstof wegneemt van hemoglobine als die zo nu en dan per ongeluk een zuurstofatoom chemisch bindt in plaats van gewoon alleen maar een beetje fysisch zoals het hoort-de zuurstof moet teder genoeg gebonden worden om aan omringende weefsels te kunnen worden afgestaan als die erom vragen. Je zou kunnen zeggen dat met al het hanteren van zuurstof, hemoglobine zich wel eens brandt en dan (methemoglobine geworden) moet worden genezen door het methemoglobine-reductase (reductie, het omgekeerde van oxydatie, is het wegnemen van zuurstof of het toevoegen van waterstofatomen); maar op den duur gaat de hemoglobine toch misschien wel aan de littekens van zijn brandwonden dood. Best mogelijk dus dat de atmosfeer binnen de rode cel gezonder is dan daarbuiten, maar mechanisch gezien is hij zeker niet rustig; het arme ding moet zijn meeste tijd de complete ronde doen: hart-grote long-arterie-long-capillair (samengeknepen worden, zuurstof opnemen, koolzuur afstaan)-vene (normale vorm terugkrijgen)-hart (klap van een hartklep oplopen, misschien, en in ieder geval wild tuimelen)-aorta (maximum snelheid)-kleinere vaten-een of andere capillair (samengeknepen worden, koolzuur opnemen, zuurstof afstaan), alles in ongeveer drieëntwintig seconden. (Tussen twee haakjes, als je toevallig een konijn bent, is die omlooptijd maar zeven-en-een-halve seconde, maar wat voor zoogdier je ongeveer ook bent, het | |
[pagina 125]
| |
kost je ongeveer zevenentwintig kloppingen van je hart om je bloed een keer helemaal rond te sturen; ook leven in het algemeen je rode cellen korter hoe kleiner zoogdier je bent. Dus misschien betekent dat: een rode cel kan maar een zeker aantal klappen van het hart, of een zeker aantal zuurstof|koolzuur-uitwisselingen verdragen.) Ook chemisch gezien kan je het leven in de erytrocyt nauwelijks loom noemen. Ik moet je wel even bekennen dat ik doodgewoon niet knap genoeg ben en op dit gebied niet genoeg weet om de juiste inlichtingen te verzamelen over alle kleine veranderingen die nodig zijn om het opnemen en afstaan van zuurstof mogelijk te maken. Als je er ook maar iets meer over wilt lezen, probeer dan vooral te beginnen met het stuk The Hemoglobin Molecule door M.F. Perutz in Scientific American, november 1964, blz. 64-76. Uit de combinatie van zijn werk met oudere en nieuwere artikelen heb ik deze indruk van al die gebeurtenissen: Neem aan dat we een bepaalde erytrocyt in de gaten houden, die op weg is naar een long. Daar, hij wordt in een longcapillair al uitgerekt tot een hoge puntmuts. Kijk nu eens naar een van die hemoglobinemoleculen in het steile deel van de muts: dat molecuul komt bijna helemaal tegen het membraan en dat membraan ligt allerdichtst tegen de capillairwand en dus vlak bij de lucht in de long. Kijk nu, desnoods, nog eens naar figuur 34. En stel je voor dat een schijf hemo aan een alfaketen (er waren vier ketens per molecuul, weet je nog wel: twee alfa en twee bèta, ieder met een hemogroep) toevallig de eerste is die door een zuurstofatoom wordt geraakt. Een watermolecuul daar vlak in de buurt, dat tot nu toe een tengere brug had gevormd via zijn twee waterstofatomen, laat onmiddellijk los, en een tweede hemogroep vangt een zuurstofatoom. Nog is het hele hemoglobinemolecuul niet merkbaar van vorm veranderd, alleen wat roder geworden. Maar nu, nog vlugger, wordt de derde hemogroep door een zuurstofatoom ontdekt, en nu wijken de twee bètaketens iets uit elkaar-daarbij geholpen door de alfa's-waardoor de vierde hemogroep op een of andere manier een paar honderd maal makkelijker voor zuurstof te vinden wordt dan hij eerst was, en daar komt dus het vierde zuurstofatoom al aan en omhelst hem. Intussen heeft het hele molecuul niet stilgezeten. Zowel het gewring van de rode cel gedurende de ruwe rit als het voortdurend gebibber dat zo typerend is voor alle warme moleculen, heeft ons rode kereltje dieper en dieper | |
[pagina 126]
| |
de cel in gedragen, waar het minder zuurstofrijke vriendjes ontmoet en snel wat uitwisselt. Hoewel dus de hele celinhoud zo dik is als een gekleurd kristal, reist zuurstof er sneller doorheen dan door helder water. Intussen heeft het hemoglobinemolecuul ook koolzuur (CO2) losgelaten, en dat heeft zich gecombineerd met een molecuul erytrocytenwater. Daardoor gebeuren allerlei ingewikkelde dingen, ionen vliegen heen en weer door het membraan en dan vangt een natriumatoom, buiten de cel gehouden of gegooid, het CO2-molecuul chemisch op. Het koolzuurmolecuul dat tenslotte in je longblaasjeslucht terechtkomt, hoeft helemaal niet hetzelfde te zijn als het molecuul dat we van de hemoglobine af zagen vallen. Intussen is de rode cel in een wijder vat geraakt en ontspant zich tot de ingedeukte schijf die hij zo graag is. Sneller en sneller door steeds wijdere vaten, draaikolken, waarachtig weer veilig het hart door, en tenslotte weer langzamer en door nauwere banen, een capillair van de linkerduim in. Houd weer eens een hemoglobinemolecuul dicht bij het celmembraan in de gaten, waar het bijna de wand van het capillair en de zuurstof-arme wereld daarbuiten kan aanraken. Het geeft een enkel zuurstof-atoom af naar die wereld, en meteen naderen zijn twee bètaketens elkaar, waardoor het om een of andere reden makkelijk wordt voor het volgende zuurstofatoom om ook van zijn hemoschoteltje los te raken. CO2 kan dan worden opgenomen, enzovoort. Als dat allemaal driehonderdduizend keer gebeurd is, beginnen de dingen in de cel te vervallen. Er is wat water weggeraakt: de cel is zwaarder van soortgelijk gewicht geworden. Zijn membraan is chemisch van gezicht veranderd, en daarbinnen in zijn lichaam kunnen een hoop dingen verkeerd zijn gegaan. Misschien hebben te veel hemoglobinemoleculen zich aan de zuurstof gebrand, zijn in methemoglobine veranderd en worden niet meer op tijd door de methemoglobine-reductase, zelf ook al een beetje ziek, gered. Het slecht onderhouden membraan wordt teder, zo teder dat het alle fut verliest: poriën gaan overal erin open, stervend zweet het hemoglobine en een hele menigte seniele eiwitmoleculen. Eindelijk in het plasma, geniet elk hemoglobinemolecuul maar een korte vrijheid; dan wordt het gegrepen door een speciaal op hem loerend eiwit: haptoglobine, en aangevallen door een troep enzymen. Die breken de alfa- en bètaketens in stukken, en de stukken in losse aminozuren. De vier hemo's, | |
[pagina 127]
| |
ieder eerst lijkend op een plat bloempje (figuur 47), worden opengemaakt, elk wordt een snoertje bloemblaadjes met fantastische kleuren: de galkleurstoffen, voor de lever om als gal te gebruiken. Uit elk hemobloempje wordt het hartje van ijzer door transferrine weggepikt en teruggebracht naar het beenmerg, voor zijn reïncarnatie in een nieuwe rode cel. 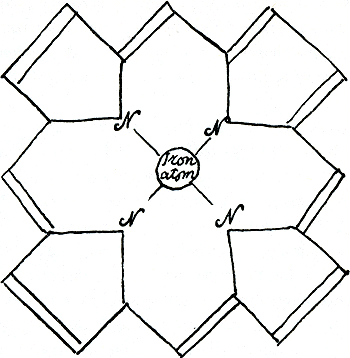
Figuur 47.
Uit sommige advertenties heb je al geleerd dat niet al het ijzer terugkeert. Je verliest wel wat, want soms moet je iets eten waar ijzer in zit. Hoe weet je wanneer? Als het zoekgeraakte ijzer ergens in een riool is weggedreven kan het je dat niet meer toeroepen; ook veel zoekrakende onderdelen van de erytrocyt zelf hebben misschien geen stem. Als je een gesloten systeem was, zou zo'n afbraakprodukt voldoende waarschuwing voor je beenmerg zijn: zoveel cellen komen van hun dood vertellen, zoveel nieuwe moeten er worden gemaakt. Maar je bent niet gesloten; de buitenwereld laat je alleen maar leven zolang je er op reageert. Als een of ander ongelukje je aan het bloeden brengt, verlies je een hoop gezonde rode cellen die voor je beenmerg geen boodschap meer kunnen achterlaten. Nee, als werkelijk het zuurstoftransport de grootste taak van de erytrocyten is, dan moet het verzoek van je buitenwereld om meer rode cellen eigenlijk een verzoek zijn om meer zuurstoftransportmiddelen, en elk weefsel dat | |
[pagina 128]
| |
bij het minste zuurstofgebrek ongerust genoeg kan worden om een of andere alarmstof te maken, een soort beenmerg-prikkelend hormoon, zou goed genoeg zijn om als rodecellen regelaar te dienen. En waarachtig, het is gebleken dat de nier zo'n weefsel is; bij gebrek aan zuurstof maakt hij een stof die erytropoietine wordt genoemd. Als je lichaam een tijd harder moet werken, of je hart wil niet hard genoeg je rode cellen rondsturen, of als je de bergen ingaat en de dunne lucht in, dan maken je nieren naar zuurstof snakkend meer erytropoietine, en al gauw verschijnen meer reticulocyten in je bloed, en de concentratie rode cellen in je bloed neemt toe tot je nieren weer gelukkig zijn. Dat is maar een deel van het hele erytrocytencontrolerende systeem. Er schijnen enige stoffen te bestaan die bepalen hoe gauw de al voltooide rode cellen uit je beenmerg kunnen worden losgelaten, het bloed in, en uit sommige proeven is op te maken dat je hersens de stroom van reticulocyten uit je merg aan en afkunnen zetten. Zo'n heel complex van opgewonden regelingen en regelingen van regelingen geeft de indruk dat het voor ons leven belangrijk is, sommige dingen (zoals zuurstoftoevoer) zo constant mogelijk te houden. Net zoals het lot van de soldaat is, te vervallen van veldslag in verveling en van verveling weer in veldslag, alleen maar om het leven van de thuiszittenden zo eentonig te houden als ooit. We moeten vol zitten met dergelijke regelende systemen; ‘negative feedback systems’ worden ze hier genoemd. Minder zuurstof-maakt meer erytropoietine -maakt meer rode cellen-maakt meer zuurstof-maakt minder erytropoietine. Te ver voorover staan-maakt reflex via ruggemergmaakt verder achterover hellen-maakt enzovoort. Daar sta je dan, trots en onomkegelbaar, hoop je. Als je te zachtjes praat zeggen je knorrige hersens dat je harder moet praten, en altijd dat je zo moet praten als je gisteren deed, en jezelf moet herkennen, zoals ook je handschrift herkenbaar en dus nogal onveranderd wil blijven. Eerlijk gezegd weet ik niet welke dingen van belang zich handhaven door constant te blijven. Voor een gelukkig huwelijk zal het wel nogal nodig zijn dat de man meer dan een paar minuten lang op zichzelf kan blijven lijken, maar wat het essentiële van hem is waarin geen verandering toelaatbaar zou zijn-daar heb ik geen idee van. Als we aannemen dat het meest algemeen constante ook het meest belangrijke is, dan behoren we tussen de verschillende waarden van | |
[pagina 129]
| |
dingen voor heel verschillende diersoorten vergelijkingen te maken. Bijvoorbeeld, geiten hebben ongeveer achttien miljoen rode cellen per kubieke millimeter en jij vijf miljoen, maar jullie hematocrieten zijn bijna hetzelfde; zij dertig, jij veertig percent. Dat komt doordat de geirecellen veel kleiner zijn dan de jouwe. De meeste zoogdieren hebben een dergelijke hematocriet; als je dus denkt dat constante waarden belangrijker zijn dan veranderlijke, dan kan je zeggen dat de doorsnede van de gemiddelde rode cel niet zo belangrijk is als de hematocriet. Dan vind je het misschien ook leuk te weten dat voor verschillende zoogdiersoorten de doorsnede van de gemiddelde rode cel meer verschilt dan de dikte, en de absolute hoeveelheid hemoglobine per rode cel meer varieert dan de concentratie (hoeveelheid per volume) in de rode cel (maar die concentratie is misschien alleen maar zo constant omdat hij nu eenmaal fysisch gesproken niet hoger kan). Wat die hematocriet betreft: best mogelijk dat een van meer dan 50% het bloed onmogelijk dik maakt; stakkertjes die door een mutatie geboren zouden worden met nieren of beenmerg die zo'n hoge waarde verlangen, zouden dan piepjong aan hartverlamming sterven. En wat de dikte van een rode cel betreft: best mogelijk dat fosfolipide-eiwit-cholesterol combinaties, die membranen moeten bouwen, alleen maar een bepaalde kromming van de erytrocytenrand kunnen maken of verdragen. Het is zelfs best mogelijk dat veel grotere structuren van dierenlichamen in het algemeen beperkt zijn tot niet al te rare grondvormen, doordat er beperkende moleculen in voorkomen die door hun moleculaire vorm zelf beperkt worden. Daarmee komen we terug op een vraag iets voor het eind van hoofdstuk ii: is de algemene gedaante van (bijvoorbeeld) hemoglobine meer belangrijk dan de atomen waaruit die is opgebouwd? Eist zijn vorm een bepaald soort vervoermiddel: de rode cel? En wordt zijn vorm zelf al bepaald door de manier waarop de hemo's moeten worden vastgehouden? Het is wel merkwaardig dat onze spieren een rode kleurstof hebben die myoglobine heet en precies zo gevormd is als de bètaketens van hemoglobine; en nog merkwaardiger dat hier en daar in het dierenrijk de gekste beesten te vinden zijn die hemoglobine maken. Andere maken weer heel andere zuurstofgrijpende pigmenten, maar alleen onder die-zoals bijvoorbeeld de worm Thalassema-welke echt hemoglobine maken, vind je de beesten die ook de | |
[pagina 130]
| |
rode cellen kunnen maken om die hemoglobine in te stoppen. Dus misschien dicteert de vorm van het molecuul werkelijk wel in wat voor vehikel het wil reizen. Als je omkijkt naar het verleden zie je niet gauw een beest zo actief als jij, dat toch in staat is alles te doen zonder rode cellen om zijn hemoglobine mee te beschermen. Er leven wel ijsvissen in de zuidpool, in water van twee graden Celsius; bijna bewegingloos liggen ze daar de dikke suspensie van levend voedsel rondom door hun geweldige bek op te zuigen. Zij hebben geen rode cellen en helemaal geen hemoglobine, hun lijven zijn wit, en de zuurstof doodgewoon fysisch opgelost in hun plasma, daar hebben ze op die manier genoeg aan. Iemand heeft uitgerekend dat wij mensen zestig maal meer dan ons gewone plasma nodig zouden hebben om daarin de hoeveelheid zuurstof in op te lossen waar onze rode cellen nu voor zorgen. Maar van al dat plasma zouden we dan driehonderd pond meer wegen. Maar om dat rond te sleuren zouden we dan drie maal zoveel zuurstof nodig hebben. Maaaar... dan moeten we toch maar dankbaar zijn voor onze rode cellen. |