
Inleiding tot de diersociologie
(1946)–Niko Tinbergen–
[pagina 133]
| |
V. De ontwikkeling van sociale structurenDe vraag, hoe sociale structuren tot stand komen, moetaen wij in twee probleemcomplexen splitsen. In de eerste plaats zien wij telkens nieuwe gemeenschappen zich vormen. Wij willen weten hoe dat in zijn werk gaat, m.a.w. hoe de ontogenese verloopt. Maar bovendien willen wij weten, hoe de gemeenschapsvormen zich in de loop der eeuwen ontwikkeld hebben, of beter hoe de soorten zich mèt hun karakteristieke manieren van gemeenschapsleven ontwikkeld hebben tot wat zij zijn, m.a.w. hoe de phylogenese verlopen is. Men weet immers sinds Darwin dat de soort niet een constant blijvend iets is, maar in de ons tegenwoordig bekende gedaante, niet meer dan een ‘momentopname’ in een ontwikkelingsreeks. Het is voor alle takken der biologie een probleem hoe die ontwikkeling der soorten zich voltrekt en waardoor hij zich zó en niet anders voltrekt resp. voltrokken heeft. De problemen zijn oorspronkelijk slechts voor wat betreft de karakteristieke vormeigenschappen der soorten in studie genomen; zodra men echter inzag dat functionele, dus ook gedragseigenschappen, even karakteristiek voor een soort kunnen zijn als vormeigenschappen, dook het probleem ook voor deze op. Wij beschouwen eerst de eerste vraag, en moeten beginnen met het onderzoeken van enkele voorbeelden. | |
1. Ontogenese.De verhouding tussen een dier, bijv. een vogel, en zijn jong ontwikkelt zich in wezen zoals de verhouding tussen een individu en een zijner organen. Aanvankelijk is het jong een eicel in het moederlichaam, een cel uit een complex, de eierstok, die men gewoonlijk een orgaan noemt. Zodra het ei bevrucht is, gaat het zich ontwikkelen, d.w.z. zich differentiëren, zich geleden. Bo- | |
[pagina 134]
| |
vendien voert het moederlichaam door een reeks ingewikkelde processen voedingsstoffen aan en vormt steunorganen en beschermende structuren, waardoor de eicel tot een min of meer afgesloten geheel, het ei, wordt. Als het ei het moederlichaam verlaat, is het dan ook al heel wat zelfstandiger dan toen het nog een eicel was; voedsel en zuurstof bijv. krijgt het nu niet meer uit het lichaam van de moeder. De zelfstandigheid is niet totaal: het ei moet door de moeder bebroed worden. Tegelijk gaat de differentiatie verder; de eicel splitst zich in cellen, die zichzelf weer splitsen; bepaalde groepen dezer cellen gaan de huid vormen, andere de darm, weer andere de hersenen, enz. Wanneer de eidop barst en het jong te voorschijn komt, worden de verhoudingen sprongsgewijs anders. De oude betrekking van het broeden blijft weliswaar, althans voorlopig, maar het eierenkeren verdwijnt en het voeren en het wegnemen van de faeces komt er bij. De jongen reageren verder op lokroepen en alarmroepen. Er blijven dus betrekkingen bestaan, al zijn deze van eenigszins subtiele aard en al zijn zij, doordat zij met behulp van de zintuigen tot stand komen, iets minder tastbaar dan de aanvankelijke, meer materieel aandoende relaties. Deze verhouding blijft, afgezien van kleine wijzigingen, bestaan totdat de jongen zich van de ouden gaan losmaken. Bij sommige soorten gebeurt dit doordat ouden en jongen alle belangstelling voor elkaar verliezen; dikwijls ook gaat de breuk van de ouden uit, die de jongen òf eenvoudig verlaten, òf hen verdrijven. Het laatste komt nogal eens bij vogels voor, wanneer zij het nest waarin de jongen nog huizen, voor een tweede broed willen gebruiken. Dikwijls ook gaat de band tussen ouderdier en jong over in de band tussen twee sociale collega's; het gezin wordt tot een troep. Deze gemeenschap was dus in zijn jonge stadia niet te onderscheiden van die tussen het lichaam als geheel en een zijner organen, en werd langzamerhand tot een tussen individu en individu, waarbij groter wordende zelfstandigheid van het ‘orgaan’ en differentiatie er van ken- | |
[pagina 135]
| |
merkend zijn. In zulke gevallen is dus een gemeenschap uit een individu ontstaan als gevolg van een verdere ontplooiïng van zijn organisatie. Deze ontplooiïng kan heel ver gaan; het hoogtepunt van een dergelijke ontwikkeling is wel bereikt in de ingewikkelde insectenstaten. Hiervan wil ik enkele voorbeelden behandelen, beginnend bij eenvoudige relaties tussen moederdier en jong en vervolgens opklimmend naar de meer ingewikkelde gevallenGa naar eind105. De meeste insectenstaten ontwikkelen zich uit een bevrucht wijfje. Het begin verschilt dus niet van een ‘gezin’. Bij vele insecten komt het niet eens tot de vorming van een echt ‘gezin’, de dieren leggen hun eieren en laten deze dan meteen in de steek. Sommige bijen- en wespensoorten echter treden in contact met hun eigen jongen. De graafwesp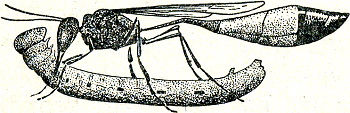 Fig. 76. De kleine rupsendoder met zijn prooi (Baerends).
Ammophila campestris bijv., de ‘kleine rupsendoder’ (fig. 76), die haar eieren in een zelfgegraven hol op een door haar verlamde rups legt, vergenoegt zich er niet zoals de meeste andere graafwespen mee, om bij elk ei dat zij legt een voorraad voedsel (rupsen) voor de larve te deponeren, maar zij brengt de larve opnieuw voedsel, wanneer deze de eerste voorraad opgegeten heeft. Wanneer de larve zich gaat inspinnen, laat zij hem echter aan zijn lot over, en zij sterft lang voordat haar kinderen uit de pop kruipen. Er zijn onder de wilde bijen enkele soorten, die in sociaal opzicht een hogere trap van ontwikkeling bereikt hebben dan deze rupsendoder. In onze streken komt een soort voor, Halictus quadricinctus, die zijn eieren op de manier der graafwespen in zelfgegraven holen in de grond legt. De uit de eieren gekomen larven voeden zich met de voorraad honing en stuifmeel, die de moeder in de cellen van het hol opgeslagen heeft. In tegenstelling tot wat bij de meeste bijensoorten geschiedt, laat de moeder het nest niet in de steek, maar maakt het uitkomen van haar kinderen mee; zij leert hen dus niet alleen als larven, maar ook als bijen | |
[pagina 136]
| |
kennen. Deze eerste jongen verlaten nu het nest niet, maar bouwen er aan verder, leggen er hun eieren en verzorgen dit nieuwe broed gezamenlijk, en wel zonder aanzien des persoons, d.w.z. zij brengen voedsel zowel voor hun eigen larven als voor die der anderen. De in de herfst uitkomende generatie is echter niet zo sociaal aangelegd en verlaat het nest, waarop ieder dier zijns weegs gaat. Deze bijen overwinteren dus solitair en elk fundeert in het volgend voorjaar een nieuw ‘gezin’. Een stap verder komen de hommelsGa naar eind106. Ook een hommel-‘staat’ wordt door één bevrucht wijfje (de koningin) gesticht. Deze koningin treedt in innig contact met haar broed doordat ze de cellen waarin de larven opgroeien af en toe opent om de voedselvoorraad bij te vullen. De eerste larven ontwikkelen zich tot wijfjes, wier eierstokken echter niet tot volledige ontwikkeling komen en die zich dan ook niet kunnen voortplanten. Deze hulpwijfjes en werksters verzorgen nu verder het broed dat uit de later door de koningin gelegde eieren opgroeit. De koningin wordt van dat ogenblik een eilegspecialiste; de werksters zorgen voor alle andere bezigheden van de familie: ze vliegen uit en halen voedsel, ze bouwen cellen, ze voederen de koningin en haar kroost. Hier treffen we dus verdeling van arbeid aan, en er is dus binnen de gemeenschap een belangrijke differentiatie der samenstellende elementen der wijfjes. In de nazomer ontstaan uit de door de koningin gelegde eieren beter ontwikkelde wijfjes en ook mannetjes. De laatsten bevruchten de wijfjes en in het najaar valt het grote gezin uiteen. Slechts de bevruchte wijfjes blijven leven. Ze overwinteren, meest apart, soms nog bij elkaar in het oude nest, maar in het voorjaar gaan ze uiteen en elk van hen gaat dan, solitair, op zoek naar een geschikt hol om daar, als koningin, een nieuw gezin te stichten. Op nog veel hogere trap van sociale ontwikkeling staan de drie statenvormende bijensoorten, waarvan de honingbij de best bekende is. In de eerste plaats gaat hier de verdeling van arbeid, zoals ik in hoofdstuk II besprak, zeer ver. In de tweede plaats echter valt de staat in het najaar | |
[pagina 137]
| |
niet uiteen, maar blijft hij, wanneer hij niet gewelddadig verstoord wordt, jaar in jaar uit bestaan. De gemeenschap overleeft dus zijn individuen, en meestal spreekt men pas in dit geval van een ‘staat’. Een nieuwe staat wordt bij de honingbijen niet door een solitaire koningin gesticht, maar door een koningin met werksters van allerlei soort. De oorspronkelijke staat, die één koningin bezit, deelt zich in tweeën even voordat er een nieuwe koningin geboren wordt; er vertrekt dan een ‘zwerm’ met de oude koningin naar een nieuwe woonplaats. Deze eerste zwerm wordt dikwijls later in het seizoen door een aantal zwermen gevolgd. die elk een jonge koningin meenemen. Nieuwe staten vormen zich hier dus, doordat de oude staten zich in twee, qua samenstelling ongeveer gelijke delen splitsen. De vergelijking met de celdeling, waarbij de twee dochtercellen gelijke erfdelen meekrijgen die ze door eigen arbeid weer moeten laten aangroeien, ligt voor de hand. Bij de mieren, waaronder men geen soorten kent die geen staten vormen, kent men eveneens twee manieren waarop een staat gesticht kan worden. Bij zeer vele soorten gaat het net als bij de hommels: een bevrucht wijfje zoekt een holte en gaat eieren leggen waaruit de eerste werksters ontstaan. Bij andere soorten echter kan de koningin niet zelfstandig leven en een nieuwe staat opbouwen, maar moet zij van het begin af aan de medewerking van een aantal werksters hebben. Bij sommige soorten wordt dit bereikt doordat de koningin met een deel der werksters een nieuwe staat sticht, bij andere soorten dringt de koningin in een vreemd nest van dezelfde soort binnen en dwingt de oorspronkelijke koningin tot vertrek. Langs deze weg van het indringen kunnen ook slavenhoudende staten ontstaan. De koninginnen van sommige soorten dringen nl. in een nest van een vreemde soort binnen, doden de volwassen mieren en verzorgen zelf het broed verder. De hieruit komende werksters blijven bij de koninging en verzorgen het van haarzelf afkomstige broed. Bij vele soorten ook huizen in één nest vele koninginnen en van tijd tot tijd vertrekt een groep werksters met een | |
[pagina 138]
| |
koningin en met een deel van het broed naar een nieuwe nestplaats.
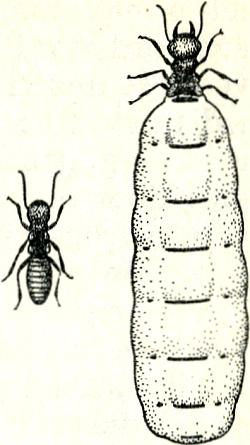 Fig. 77. Een termietenkoningspaar; het wijfje is een grote ‘elerfabriek’.
De staten der termieten zijn, zoals ik al eerder beschreef, niet zuiver matriarchaal, maar mannetjes en wijfjes dragen aan het geheel evenveel bij; er is een ‘koningspaar’ (fig. 77), en de ‘werkster’-kaste bestaat uit evenveel mannetjes als wijfjes. Op een gegeven ogenblik ontstaan in een termietenstaat geslachtsdieren, en wel mannetjes en wijfjes in gelijk aantal, en deze zwermen tegelijk uit. Na het zwermen verliezen zij de vleugels en op de grond vindt de paarvorming plaats, doordat de wijfjes met behulp van bepaalde geurklieren de mannetjes lokken. De dieren van zulk een paar zijn nog niet geheel geslachtsrijp; zij komen voorlopig nog niet tot paren, maar gaan samen een nestholte graven, het begin van een termietenheuvel, de woonplaats van het toekomstige volk. Na enige tijd rijpen de dieren en pas dan voltrekt zich de paring en het afzetten van eieren. De larven der termieten zijn lang niet zo hulpbehoevend als die der bijen en mieren, en nemen al spoedig deel aan de werkzaamheden. Door een geleidelijke ontwikkeling worden zij tot de volwassen arbeiders en arbeidsters van de verschillende ‘kasten’. In alle besproken gevallen zien wij een gemeenschap geleidelijk groeien uit de verhouding moederdier-eicel, of ouders-ei. De ontstaanswijze van zulke gemeenschappen zou men dus met ‘groei’, of ‘ontplooiïng’ kunnen betitelen.
Geheel anders, ja op het eerste gezicht tegengesteld hieraan, is de ontwikkeling van de gemeenschappen die zich vormen door aaneensluiten van reeds zelfstandig bestaande individuen. Dit gebeurt bijv. wanneer twee dieren een paar gaan vormen. In de voorgaande hoofdstukken | |
[pagina 139]
| |
heb ik hiervan vele voorbeelden behandeld. Min of meer plotseling ontstaan hierbij banden tussen de individuen die er tevoren niet waren, en die hun samenwerking in dienst van het grotere geheel verzekeren. Dit is dus een wijze van ontstaan van een gemeenschap die men als ‘constructie’ zou kunnen betitelen.
Hoe ontwikkelen zich nu de betrekkingen tussen de individuen bij het ontstaan van een nieuwe gemeenschap? Wij hebben gezien, dat de meeste betrekkingen berusten op wat wij kortheidshalve als aangeboren gevoeligheden en aangeboren handelingen betitelen. Het goed functionneren van deze aangeboren gedragselementen wordt meestal verzekerd doordat zij hun ontwikkeling, hun groei, voltooid hebben vóórdat zij in functie hoeven te treden. Een vogel is gevoelig voor de aanblik van een ei van zijn soort, en voelt bij die aanblik een drang in zich opwellen om het te bebroeden, al voordat hij zijn eerste eigen ei gelegd heeft. De drang tot jongen voeren bestaat al voordat de eigen jongen geboren zijn. Ook bij de andere wijze van gemeenschapsvorming, het aaneensluiten van individuen op de constructieve manier, liggen de potenties tot ageren en reageren van tevoren klaar. Noch een heivlinder, noch een stekelbaars, noch een blauwe reiger, om maar enkele voorbeelden te noemen, behoeven, zoals wij gezien hebben, te leren hoe zij hun geslachtspartner te herkennen en te bejegenen hebben. Zowel dus bij de ontplooiïng van een gemeenschap als bij de constructie van een gemeenschap komt bij de verandering der constellatie het nieuwe verband tot stand door het in werking treden van potenties die al van tevoren gereed lagen.
Met het vestigen van een band tussen twee individuen is echter de ontwikkeling van die band nog niet voltooid. In de eerste plaats kan hij in de loop der tijd intensiever worden of verzwakken, dus gradueel veranderen. De zorgen van een roodkaak voor zijn eieren bijv. veranderen | |
[pagina 140]
| |
ietwat in de loop van de ontwikkelingstijd der eieren. Als deze nog jong zijn, voert hij hun betrekkelijk weinig vers (zuurstofrijk en koolzuurarm) water toe. Hij doet dit door een waterstroom op te wekken met behulp van zijn vinnen, wat als ‘waaieren’ betiteld wordt. Naarmate zich nu de eieren ontwikkelen, nemen duur en frequentie van het waaieren toe. Dit gebeurt ook wanneer men ev. van de eieren uitgaande prikkels, die hiervoor aansprakelijk zouden zijn, uitschakelt, bijv. door de eieren van tijd tot tijd door verse, jongere eieren te vervangen. Er ontwikkelt zich dus iets in de roodkaak zelf, dat aansprakelijk moet zijn voor de graduele verandering in zijn gedrag. Op dergelijke wijze ontwikkelt zich de broeddrang bij een broedende vogel. Naarmate de ontwikkeling van het ei voortschreidt, wordt de broeddrang sterker. Dit gebeurt ook in gevallen waar de eieren dood zijn; ook hier hebben wij dus met een ontwikkeling in de oude vogel te maken. Ingrijpende processen treden op onder invloed van leerprocessen. Door leerprocessen worden de betrekkingen dikwijls hechter in die zin, dat zij specifieker worden, dat de reactor in de loop van de tijd minder ontvankelijk voor signalen van andere dan de bij hem behorende actor wordt. In de regel merkt men zulke veranderingen alleen wanneer afwijkende omstandigheden optreden. Door leerprocessen komen bijv. alle persoonlijke bindingen tot stand die in hoofdstuk III besproken zijn. De meeste dier persoonlijke bindingen komen waarschijnlijk langzamerhand en gradueel tot stand. Daarop wijst bijv. het feit dat vogels, waarvan bekend is, dat zij hun jongen persoonlijk kennen, hiervan pas blijk geven als deze enige dagen oud zijn. Het heeft er alle schijn van, dat zij deze dagen nodig hebben. Er komt echter ook een soort van specifieker worden van de relaties voor, die zich van deze langzame leerprocessen onderscheidt, doordat zij in heel korte tijd haarbeslag krijgt. Wanneer men bijv. een ei van de grauwe gans in de broedstoof laat uitkomen, en het jong, zodra het uitgekomen is, onder zijn hoede neemt zonder het met | |
[pagina 141]
| |
de eigen soort te confronteren, wordt men in een minimum van tijd in de ogen van dat kuiken tot de bij hem behorende oudervogel. Het is onmogelijk, zulk een jong bij ganzen onder te brengen; het herkent hen niet meer als soortgenoot en loopt luidt piepend naar de menselijke pleegvader. Een kuiken echter dat onder een gans uitgebroeid wordt, sluit zich zonder meer bij de oude ganzen aan en wil van de mens niets weten. Het dier sluit zich dus aan bij de eerste levende wezens die het na het uitkomen ontmoet, en beschouwt vanaf dit ogenblik alleen die wezens als zijn ouders. Het merkwaardige is nu, dat deze connectie in bijzonder korte tijd tot stand komt. Volgens Heinroth moet men een ganzenkuiken dat in de broedstoof uitgekomen is en dat men bij ganzenouders wil onderbrengen, dadelijk nadat men de broedstoof geopend heeft, in de zak steken en het zó naar zijn as. pleegouders brengen, om te voorkomen, dat het zich aan de mens hecht. Het lijkt dus, of zulk een pasgeboren kuiken zich in enkele seconden het uiterlijk van zijn pleegouders inprent. We hebben dus met een ongeëvenaard snel leerproces te doen. Sommige onderzoekers menen dat dit zich-inprenten (‘Prägung’) zich bovendien van andere soorten van leren onderscheidt doordat het dier deze indrukken niet vergeten kan. Er zou als het ware in deze eerste seconden iets gefixeerd worden, en deze fixatie zou niet ongedaan gemaakt kunnen worden. Dit is echter nog niet strikt bewezen. Zulk een jonge gans wordt dus geboren met een zeer weinig bepaalde gevoeligheid voor signalen die van de ouders uitgaan. Vóór de determinatie volgt hij alles wat zich beweegt. Trekt men aan zulk een ‘onbeschreven’ kuikentje een kussen voorbij, dan volgt het dit. Men heeft eens jonge bergeenden waargenomen, die hardnekkig een roeibootje volgden. Bij dit eerste volgen nu bindt het dier zich aan zijn leider. Hoe ingewikkeld deze verschijnselen zijn, blijkt wanneer men nu de gevoeligheid van zulke op de mens ingestelde, ‘zich als mens beschouwende’ ganzen voor de signalen | |
[pagina 142]
| |
van de soort verder onderzoekt. Dan blijkt bijv. dat zij ook in hun menselijken vader die signaalbeweging verwachten, begrijpen en opvolgen, die oude ganzen in de regel uitvoeren. Laat men bijv. zulke ganzen, als ze opgegroeid zijn, vrij rondvliegen, dan kan men hen tot neerstrijken bewegen door hard voor hen uit te lopen en, als zij in de buurt gekomen zijn, plotseling plat op de grond te vallen, zodoende de beweging van het neerstrijken uitvoerende zo goed als een mens dit maar kanGa naar eind107. Ondanks hun omstelling op mensen behouden deze dieren dus de aangeboren gevoeligheid voor dit signaal van hun eigen soort. Men weet niet, of dit merkwaardige leerproces ook bij andere dieren voorkomt. Wèl weet men dat er vogelsoorten zijn, waarvan de jongen hun eigen soort wel degelijk kennen zodra ze uit het ei kruipen, en die men dus niet kunstmatig aan de mens kan koppelen. Een wulpenjong bijv. beschikt over een dergelijke fijne aangeboren gevoeligheid. Veel verder reikt onze kennis echter nog niet en ook hier staat het onderzoek dus nog in de kinderschoenen. Enkele aanwijzingen dat het zich-inprenten ook bij andere dieren voorkomt, heeft men wel en wel bij vissen. De al meer genoemde Cichlide Hemichromus bimaculatus bijv. leert het uiterlijk van de jongen van zijn soort kennen (leert de verzorgende handelingen te beperken tot jongen van zijn eigen soort) bij de eerste keer dat hij jongen heeftGa naar eind108. Verwisselt men de eieren van een jong paartje, dat voor het eerst eieren gelegd heeft, met eieren van een andere soort, dan verzorgt het paar de hieruit komende jongen goed. Doet men hetzelfde bij een ervaren paar, dat al eens of meermalen jongen opgevoed heeft, dan eet het de vreemdsoortige jongen op. Legt het eerstgenoemde paar nu echter later weer eieren en laat men hen deze houden, zodat zij nu met jongen van hun eigen soort geconfronteerd worden, dan eten zij hun eigen jongen op! Ze erkennen namelijk na hun eerste ervaring alleen nog maar de jongen van de hun oorspronkelijk ondergeschoven soort. Hoe snel hier de fixatie in zijn werk gaat en wanneer zij precies plaats vindt, is echter niet bekend. | |
[pagina 143]
| |
Er is aan dit leren-kennen van de soortgenoot, of het nu vergelijkbaar is met het behandelde ‘zich-inprenten’ of niet, nog één interessant aspect. Wanneer men een jong kauwtje uit het nest neemt en thuis opvoedt, dan leert het zijn menselijke pleegvader als zijn eigen vader te beschouwen, d.w.z. hij is niet schuw voor hem, zoekt hem integendeel op en bedelt bij hem om voedsel. Wanneer zulk een mensenkauwtje begint te vliegen, bevredigt menselijk gezelschap hem niet meer en hij zoekt aansluiting bij dieren, die eveneens vliegen. Voor alles wat hij vliegend doet verlaat hij de mens. Zijn er wilde kauwen of kraaien in de buurt, dan worden die zijn vliegmakkers. Samen met hen gaat hij naar hun voedselgebieden en blijft daar het grootste deel van de dag in hun gezelschap. Wordt hij nu geslachtsrijp en komt hij, een jaar oud, in bronst, dan blijkt dat hij door zijn opvoeding onder mensen, en ondanks het feit dat kraaien zijn vliegmakkers zijn, zichzelf tòch in zeker opzicht als mens is gaan beschouwen: hij pronkt en baltst tegen een of ander menselijk wezen in zijn omgeving. Hij beschouwt zich daarentegen weer als kauwtje als zijn ouderlijke instincten ontwaken en hij de lust in zich voelt opkomen om jongen te voeren. Deze drang viert hij namelijk alleen op jonge kauwtjes bot. Afhankelijk van de aard van zijn bezigheden richt zulk een dier zich dus nu eens tot mensen, dan weer tot kauwtjes, dan weer tot andere vogels. Van één kauw, die een zekere vermaardheid verworven heeft, Prof. Lorenz' ‘Tschock’, is bekend dat hij zijn pleegvader als vader of moeder beschouwde, bonte kraaien als sociale genoten, een jong meisje als zijn vrouw, en een jong kauwtje als zijn kind. Deze dieren zien hun omgeving, speciaal de soortgenoot, dus wel op een eigenaardige, verbrokkelde manier. Zij leren niet, zoals we zouden veronderstellen: zó ziet mijn soortgenoot er uit, om dan al hun sociale gedragingen er op te richten, maar afhankelijk van de aard van hun bezigheden van het ogenblik herkennen ze een soortgenoot nu eens aan het ene kenmerk, dan weer aan andere kenmer- | |
[pagina 144]
| |
ken. Bij elke bepaalde instinctieve instelling (ouderlijke stemming, sexuele stemming enz.) behoort een bepaalde gevoeligheid. Op grond van die, oorspronkelijk dikwijls heel vage, gevoeligheid, sluit het dier zich tot een gemeenschap met andere dieren aaneen, die bij deze gevoeligheden passen, of om het anders te zeggen: die hem op grond van zijn vage besef aanstaan. Tijdens dat aansluiten prent hij zich dan het uiterlijk van zijn kameraden precieser in; bij het kauwtje komt het dan zelfs tot een persoonlijk herkennen. Het is duidelijk dat een kauwtje zich zulke uiteenlopende kameraden alleen kiest, wanneer hij in abnormale omgeving opgevoed is. In de natuur bindt hij zich aan kauwtjes doordat hij door kauwtjes opgevoed wordt, bij kauwtjes is als hij gaat vliegen en jonge kauwtjes uit zijn eieren ziet komen, kortom doordat hij telkens als het ogenblik van zich-inprenten komt, alleen kauwtjes om zich heen heeft. Uit de waarnemingen over de ‘mensenkauwtjes’ blijkt echter, dat een kauwtje geen beeld van ‘de soortgenoot’ heeft, maar het kent slechts ‘een makker om voedsel aan te vragen’, ‘een makker om mee uit te gaan’, ‘een makker om mee te trouwen’ en ‘een makker om te voeren’. De verschillende makkers moeten aan verschillende eisen voldoen, maar al die eisen kloppen precies met wat een echte kauw biedt. Bij de verschillende handelingsrichtingen ziet een kauw echter verschillende aspecten van zijn soortgenoot. En misschien is het voor een kauwtje wel even moeilijk of liever onmogelijk, om zich het abstracte beeld van een soortgenoot in het algemeen te vormen, als het voor ons onmogelijk is, om uit de verschillende aspecten waaronder wij de natuur zien, zoals het natuurwetenschappelijke en het religieuze, een algemeen natuurbeeld te abstraheren. Aan welke kenmerken nu precies een dier voldoen moet, om ‘sociale kameraad’, ‘sexuele kameraad’ e.d. te worden, waardóór dus de door mensen opgevoede kauw nu juist een meisje als vrouw nam, en bonte kraaien als sociale kameraden, moet nog onderzocht worden. Men zou | |
[pagina 145]
| |
hiervoor de bijzondere gevoeligheid van de soort nauwkeurig moeten onderzoeken. Ook hier staan we dus aan het begin van de uitvoering van een programma, en niet nabij de oplossing onzer problemen. | |
2. Phylogenese.Beschouwen wij thans het tweede probleem, dat van de ontwikkeling der levensvormen in de loop der eeuwen en wel meer in het bizonder: hoe hebben de sociale soorten zich ontwikkeld? Het algemene probleem van de ontwikkeling der levensvormen heeft men bij de studie van de bouw der dieren reeds lang geleden in studie genomen. Eerst betrekkelijk kort geleden heeft men de vraag ook op de gedragingen der dieren toegepast. Men staat hier echter voor grotere moeilijkheden en wel hoofdzakelijk doordat directe historische gegevens ontbreken. Over de bouw der dieren uit vroegere epochen lichten de fossielen ons betrekkelijk goed in; maar fossielen bewegen zich niet, en hoe de gedragingen en het gemeenschapsleven dezer uitgestorven dieren geweest zijn, kunnen zij ons dus niet vertellen. Enkele, overigens onbelangrijke, uitzonderingen zijn er weliswaar. De skeletten der in historische tijd gestorven reuzenduiven van Martinique en Rodriguez dragen op de vleugels grote beenknobbels, die ongetwijfeld als ‘sporen’ in dienst gestaan hebben van het vechten met de vleugel. Wij kunnen ons dus enigszins voorstellen, hoe deze duifachtige vogels gevochten moeten hebben. Zulke uiterst fragmentarische gegevens kunnen ons echter niet zo heel veel begrip bijbrengen van de sociale structuur dezer dieren; de palaeontologie valt hier als hulpwetenschap dus helemaal uit, en dat betekent een grote handicap. De kans, om in de korte periode waarin wij zelf ooggetuige zijn van het gebeuren op het wereldtoneel, enige verandering in de sociale structuur van een soort te zien optreden, is wel heel gering. Men heeft wel eens kleine sprongsgewijze veranderingen in de bouw van een soort zien optreden, bijv. in de vlinder Amphidasis betularia L.Ga naar eind109. In de populatie van deze schorskleurige vlindersoort | |
[pagina 146]
| |
traden omstreeks 1850 plotseling enkele donkere individuen op. In de loop der jaren werd het aantal dezer donkere vlinders steeds groter, en tenslotte vormden zij de overgrote meerderheid der individuen. De soort veranderde hier dus in korte tijd van kleur. Dergelijke gevallen heeft men op sociologisch gebied echter nog nooit waargenomen; zij zouden trouwens heel moeilijk vast te stellen zijn. Als enige bruikbare, zij het dan ook weinig directe, methode blijft ons daarom die van de onderlinge vergelijking der thans levende soorten over. Daar men hiermee in de sociologie nog niet heel ver gevorderd is, volsta ik met bij wijze van illustratie enkele voorbeelden te geven, om de gedachtengang duidelijk te maken. Het onderzoek van de ontwikkeling van de bouw der dieren heeft aangetoond, dat de dieren die men gewoonlijk als de ‘lagere’ dieren beschouwt, in het algemeen meer gelijkenis met dieren van vroegere perioden vertonen dan de ‘hogere’ dieren. Met andere woorden, dieren van het bouwplan der zoogdieren of vogels zijn over het algemeen jonger dan dieren van het bouwplan van bijv. slakken en zeesterren. Dat wil natuurlijk niet zeggen, dat de tegenwoordig levende vogels ‘jonger’ zouden zijn dan de tegenwoordig levende zeesterren; allen zijn immers even jong. Alleen de tegenwoordige zeesterren hebben zich minder ver van hun oorspronkelijke bouwplan verwijderd dan de tegenwoordige vogels. Door na te gaan in welke opzichten de hogere dieren sociologisch van de lagere verschillen, kan men dus enig idee er van krijgen, welke vormen van gemeenschapsleven ‘modern’ zijn. Deze algemene vergelijking leidt inderdaad tot enige, zij het ook ietwat vage conclusies. Aangezien de sexuele gemeenschapsvorm bij de laagste dieren voorkomt die men kent, de eencelligen, neemt men aan, dat deze gemeenschapsvorm zeer oud moet zijn. Bij vele eencelligen is echter het onderscheid tussen een ‘mannetje’ en een ‘vrouwtje’ zeer veel geringer dan bij | |
[pagina 147]
| |
de hogere dieren. De verregaande arbeidsverdeling tussen mannetje en vrouwtje is dus wel pas opgetreden na een periode waarin mannetje en wijfje weinig of niet van elkaar verschilden. De sociale verhoudingen in engere zin kent men ook reeds bij eencelligen. Zodra het sociale verband bestaat uit een materieel aaneenhechten zoals bij Volvox e.a., aarzelt men, zoals in hoofdstuk IV uiteengezet werd, om van sociale gemeenschap te spreken en gebruikt men, zij het ook schoorvoetend, de term individu. Bij veelcellige dieren treft men ook in de lagere stammen sociale aanleg aan. Relaties tussen ouderdier en kind zijn een betrekkelijk moderne vorm van gemeenschapsleven, want men treft hen bij lagere dieren niet aan. In de regel wordt na het leggen der eieren het contact verbroken. Verder dan uitspraken van dit kaliber brengt ons dit soort van vergelijking niet. Wèl kan men nog zeggen, dat, aangezien de hogere dieren over betere zintuigen beschikken dan lagere, en zij dus de buitenwereld gedétailleerder waarnemen, hun gemeenschapsleven ingewikkelder, hun relaties hechter en veelzijdiger en verder strekkend zijn. De afstand waarop vogels elkaars gedragingen kunnen beïnvloeden, de wijze waarop zij zich bij elkaars bewegingen kunnen aanpassen, verschillen hemelsbreed van wat slakken in dit opzicht presteren. Daar komt nog bij, dat bij de hogere dieren meer dan bij lagere zgn. hogere geestelijke vermogens in het spel komen, o.a. verschillende vormen van leervermogen en alles wat daarmee samenhangt.
Men kan echter ook op een andere manier vergelijken. In plaats van de grote stammen als geheel onderling te vergelijken kan men ook binnen de stam kleinere groepen, ja soorten met elkaar vergelijken en dan gaat men weer andere, minder omvattende, meer gedétailleerde problemen zien. Vergelijkt men bijv., zoals hierboven al is gebeurd, de vele bijensoorten met elkaar, dan merkt men dat het merendeel dier soorten eenzaam leeft en dat er maar drie | |
[pagina 148]
| |
statenvormende soorten zijn. En uit het gegeven overzicht blijkt wel al, dat er tussen de sociale en de solitaire bijen ‘tussenvormen’ voorkomen. Daar verreweg de meeste bijen niet-statenvormend zijn, mag men wel aannemen, dat deze toestand de oorspronkelijke is en dat statenvorming een betrekkelijk moderne topprestatie van deze groep is, en niet juist de oude, primitieve toestand. Aangenomen dus, dat de ontwikkeling hier gegaan is van solitair naar sociaal en niet omgekeerd, kunnen wij met grote mate van waarschijnlijkheid concluderen dat de ‘tussenvormen’ ongeveer de weg aangeven waarlangs de ontwikkeling gegaan moet zijn. Ongeveer, want we kunnen ons niet voorstellen, dat deze ‘tussenvormen’ werkelijk op een lagere trap van ontwikkeling zijn blijven staan. Zij hebben hun sociale gewoonten wellicht later ontwikkeld, of langzamer. Zij kunnen zich in iets andere richting dan de honingbij ontwikkeld hebben. Maar daar al die primitievere sociale soorten als gezin van moeder met kinderen ontstaan, moeten wij wel aannemen, dat de staten der honingbijen zich ook uit het gezinsverband ontwikkeld hebben, d.w.z. uit de gemeenschap van moeder met kinderen. Bij de andere statenvormende insecten, nl. de mieren en de termieten, helpt vergelijking ons minder. Wij kennen nl. alléén statenvormende en geen solitaire mieren of termieten. Toch valt er nauwelijks aan te twijfelen, gezien de structuur der eenvoudigste staten dezer groepen, dat ook hier in het gezin de oorsprong van de staat gezocht moet worden. Maar terwijl bij de mieren de mannetjes even goed als bij de bijen ondergeschikte, slechts tot paren geboren, leden van de maatschappij zijn, zijn zij bij de termieten even volwaardig als de wijfjes; alle ‘kasten’ bestaan hier uit evenveel mannetjes als wijfjes.
Men kan de vergelijking tot nog minder omvattende verschijnselen beperken, en van vergelijking der sociale structuur als geheel afdalen tot vergelijking van zijn onderdelen. Dit heeft men met enig succes gedaan in het | |
[pagina 149]
| |
geval der signaalbewegingen en wel in het bizonder der oversprongbewegingen. In hoofdstuk III zagen wij, dat oversprongbewegingen sterk lijken op bewegingen uit het dagelijks leven. Men kan dan ook spreken van oversprong-eten, oversprong-verenpoetsen, oversprong-slapen enz. De gelijkenis is echter geen identiteit; er is verschil tussen oversprong-eten en echt eten, tussen de oversprongbeweging en zijn ‘voorbeeld’. De dreigbeweging van de stekelbaars is oversprong-eten, maar het is geen volledige eetbeweging: het ophappen, fijnkauwen en doorslikken van prooi blijft achterwege. Zo is het met vele, zo niet alle oversprongbewegingen: zij zijn brokstukken van handelingen, gedeelten van hun ‘voorbeelden’. Sommige oversprongbewegingen wijken in nog andere opzichten van hun voorbeeld af. Om bij het dreigen van de stekelbaars te blijven: hierbij zet het dier een buikstekel, soms beide tegelijk, op en keert den tegenstander zijn zijkant toe, hem zo de dreigende stekel en tevens de rode onderkant demonstrerend. De beweging wordt als dreigbeweging als het ware onderstreept door de wapens te tonen en door hem opvallender te maken dan de eetbeweging zelf, nl. door den tegenstander steeds de rode kleur te tonen, die immers intimiderend werkt op een indringer. Bij vele oversprongbewegingen die signaalfunctie hebben, kent men dat verschijnsel: de beweging wijkt zodanig van zijn voorbeeld af, dat hij opvallender wordt. Men zegt: de beweging is ‘geritualiseerd’, tot speciale ritus geworden. In deze uitdrukkingswijze ligt al opgesloten dat men zulk een signaalbeweging als uitvloeisel van een ontwikkeling ziet. Hiertoe noopt juist de vergelijking. Bij verschillende nauw verwante diersoorten vindt men namelijk dikwijls dezelfde oversprongbeweging in verschillende graad van ritualisatie. Men kan dikwijls de verschillende soorten rangschikken, waarin aan het ene einde soorten staan waarbij de bewuste oversprongbeweging slechts weinig van zijn voorbeeld afwijkt, aan het andere einde zulke | |
[pagina 150]
| |
 Fig. 78. ‘Oversprongpoetsen’ bij vier eendensoorten (Lorenz).
waar die afwijking juist groot is. Zo hebben verschillende kraanvogelsoorten een dreigbeweging die, evenals dat bijv. bij de spreeuw het geval is, oversprong-verenpoetsen is. Bij de gewone kraanvogel wijkt het oversprong-verenpoetsen zó weinig van het het echte verenpoetsen af, dat iedere waarnemer het dadelijk als verenpoetsen herkent. Bij andere kraanvogels wijkt het sterker daarvan af, en er is een soort waarbij men de dreigbeweging nooit als verenpoetsen zou herkennen, wanneer men de overeenkomst met de soorten niet zag, waar men de beweging ‘nog juist’ wèl als oversprongpoetsen kan herkennen. Bij eenden ziet men iets dergelijks; hier komt oversprongpoetsen als bestanddeel van de balts voor (fig. 78). Ook hier zijn soorten waar het weinig van zijn voorbeeld afwijkt (gewone wilde eend) en soorten waar het amper als poetsbeweging herkend kan worden (mandarijneend)Ga naar eind110. In de vormleer gaat men trouwens op precies dezelfde wijze te werk. Als voorbeeld neem ik de ‘schaar’ der kreeftenGa naar eind111. Een hommer, de kreeftensoort die men in de zeeviswinkels wel voor de ramen ziet liggen, heeft zoals bekend vier paar looppoten en draagt daarvóór een formidabele schaar. Men neemt aan, dat die schaar zich door specialisatie uit een looppoot ontwikkeld heeft, dus dat er | |
[pagina 151]
| |
vijf paar looppoten zijn, waarvan de voorste tot schaar is geworden. Hiervoor gebruikt men (onder andere) het volgende argument: er zijn kreeftensoorten, die vijf paar looppoten bezitten, en waarbij het eerste paar heel weinig van de volgende paren afwijkt. Er zijn ook soorten, wier voorste pootpaar duidelijk van de overige verschilt, hoewel de overeenkomst in bouw toch groot is. Bij weer andere soorten is het verschil zó groot, dat men al neiging heeft, de voorste poot als ‘schaar’ te betitelen. En bij de hommer ziet men de overeenkomst tussen schaar en looppoot alléén als men de genoemde tussenvormen eveneens kent. Men ‘herkent’ dus de schaar als poot, als men de verschillende soorten in een reeks rangschikt, waarin aan het ene einde de dieren met een weinig afwijkende eerste poot, aan het andere einde die met een gespecialiseerde schaar zet. Men kan deze reeks grafisch door twee divergerende lijnen voorstellen; de basislijn geeft de gewone poten aan; de andere lijn, die zich steeds verder van de basislijn verwijdert, de eerste poot. De soorten zijn aangegeven door vertikale lijnen. Hoe verder het snijpunt van de soortslijn met de twee lijnen uiteenliggen, hoe groter het verschil tussen de schaar en zijn ‘voorbeeld’ is. Hetzelfde diagram zou men van de genoemde oversprongbewegingen kunnen maken. In plaats van ‘schaar en zijn voorbeeld’ behoeft men slecht te zeggen ‘oversprongbeweging en zijn voorbeeld’. Nu kan men zulk een reeks, die alléén op de vergelijking der thans levende vormen berust, altijd in twee richtingen lezen. Dat wil zeggen, men kan aannemen, dat de ontwikkeling, waarvan wij immers altijd een statische momentopname zien, zich in de richting van een grotere divergentie, van een sterke ritualisatie beweegt, òf omgekeerd, dat er een tendentie tot verminderen der ritualisatie werkzaam is. Wat betreft de vormeigenschappen kan men de richting der ontwikkeling vaststellen door onderzoek der fossiele vormen en dikwijls door onderzoek der individuele ontwikkeling, want het embryo doorloopt dikwijls ontwikkelingsstadia die in sommige opzichten op de voorouders gelijken. | |
[pagina 152]
| |
Waarom de palaeontologie in ons geval van geen waarde is, zette ik hierboven uiteen. De embryologie kan ons echter in de sociologie al evenmin helpen, wat onder andere daaraan te wijten is, dat een signaalbeweging in de individuele ontwikkeling niet optreedt vóórdat hij nodig is, en dàn blijkt hij kant en klaar te zijn. De veronderstelling, dat de ontwikkeling op vermindering der ritualisatie gericht is, wordt zeer onwaarschijnlijk wanneer men zich realiseert, dat ritualisatie functie heeft, dat zij een aanpassing is. Het is nl. gebleken, dat een oversprongbeweging die sterk op zijn voorbeeld lijkt, als signaal wel eens misverstaan wordt. De volgende waarneming legt hiervan sprekend getuigenis af. De dreigbeweging waarmee een zilvermeeuw indringers in zijn territorium bejegent, is een oversprong-nestelbeweging: net als bij het ‘kuiltjedraaien’ drukt hij de borst in de grond en trappelt met de poten. Wanneer deze beweging als dreigement gebruikt wordt, gaat hij gepaard met bewegingen die aanvalsdrang uitdrukken: kop- en nekhouding en de driftigheid der beweging maken het de menselijke waarnemer in vele gevallen mogelijk om uit te maken of het dier agressief gestemd is, of echt wil nestelen. Zó gering is echter de ritualisatie, dat men zich soms toch in de betekenis vergist. Datzelfde overkomt nu blijkbaar een zilvermeeuw ook wel eens. Het echte nestelen heeft eveneens signaalfunctie, maar met een heel andere uitwerking dan het oversprongnestelen. Een kuiltjedraaiende meeuw noopt door zijn gedrag zijn maat juist tot meenestelen. Men is nu eens in een zilvermeeuwenkolonie getuige geweest van het volgende voorval. Een door de kolonie zwervend, vermoedelijk ongepaard, mannetje kwam het hof maken aan een gepaard wijfje, dat op dat ogenblik alleen op haar territorium was. Zij moest van de avances van de vreemdeling niets hebben en ging als reactie dreigen, d.w.z. ze liep een eindje in agressieve houding op het mannetje toe en ging, nog op meters afstand van hem, zitten nestelen. De man vatte dit verkeerd op en rende opeens op haar | |
[pagina 153]
| |
af en maakte aanstalten om ‘gezellig mee te nestelen’. Dit was nu juist niet wat het wijfje bedoeld had, en zij viel hem dan ook prompt aan. Hier deed zich dus het merkwaardige geval voor, dat een zilvermeeuw het oversprongnestelen ten onrechte als echt nestelen opvatte. De veronderstelling ligt voor de hand, dat dit niet gebeurd zou zijn, wanneer oversprongnestelen en ‘voorbeeld’ niet zo sterk op elkaar leken, m.a.w. wanneer de oversprongbeweging sterker geritualiseerd zou zijn geweest. Wij moeten dus wel aannemen, dat een signaalbeweging des te beter werkt, naarmate hij sterker geritualiseerd is. Ritualisatie is dus functionele specialisatie en is van voordeel voor de gemeenschap, en dit geeft ons zeker het recht te concluderen, dat de ontwikkeling gericht is op divergentie, op vergroten der verschillen, op versterken der ritualisatie. Het loont de moeite, om verder na te gaan, in welk opzicht de geritualiseerde oversprongbewegingen van hun voorbeelden afwijken, om te zien of ritualisatie soms bepaalde wetten volgt. En dan blijkt, dat geritualiseerde oversprongbewegingen zich van niet of weinig geritualiseerde oversprongbewegingen juist in die kenmerken onderscheiden, die wij boven (hoofdstuk III) als karakteristiek voor signalen in het algemeen hebben leren kennen: opvallendheid, eenvoud, specificiteit. Het oversprongpoetsen van de wilde eend-woerd, dat als signaal bij de balts dienst doet, bestaat uit één enkele, maar heel opvallende ruk van de snavel langs de schachten der slagpennen. Hierbij wordt de vleugel zó opgetild dat de blauwe spiegel goed zichtbaar wordt; bovendien wordt de beweging nog opvallend gemaakt door het geluid dat bij het krassen van de snavel over de schachten van de slagpennen ontstaat. Vergelijkt men het oversprongpoetsen bij verschillende eenden onderling, dan blijkt heel duidelijk, dat bepaald gekleurde structuren in dienst staan juist van deze signaalbeweging. Het sterkst is dat wel bij de mandarijneend, wiens poetsen tot het aantippen van één enkele slagpen geritualiseerd is, een veer die zich, in afwijking van zijn | |
[pagina 154]
| |
buren de gewone slagpennen, tot een reusachtige oranje vlag heeft ontwikkeld. Ritualisatie is dus het ontwikkelen van signaalkenmerken en wel met behulp van beweging, vorm èn kleur. Het lijkt er in zulke gevallen dus erg op, of de ontwikkeling van zulk een signaal begint bij de beweging en zich dan uitbreidt over vorm en kleur, waarbij bepaalde structuren als het ware in dienst van de signaalbeweging gesteld worden. In dezelfde richting wijst ook het onderzoek van signaalbewegingen van andere oorsprong. Oók intentiebewegingen en óók ontwakende handelingen kunnen in geritualiseerde vorm als signaal dienst doen (vgl. hoofdstuk III). Hoewel hierover nog slechts weinige onderzoekingen verricht zijn, schijnt het toch dat deze drie soorten van signaalbewegingen alle op dezelfde wijze aan ritualisatie onderworpen zijn, waardoor zij een tendentie hebben, zich tot steeds fantastischer vormen te ontwikkelen. Nu gelden deze conclusies niet voor alle signaalbewegingen, ja, slechts een klein gedeelte der bekende signaalbewegingen kan men tot een dezer categoriën terugbrengen. Een groot aantal heeft, althans op het eerste gezicht, niets met een der drie te maken. Althans op het eerste gezicht. Want nu wij weten, dat een op zichzelf ‘onherkenbare’ beweging als de dreigbeweging van de Mantschoerijse kraan door vergelijkende studie als sterk geritualiseerde oversprongbeweging herkend werd, nu is het duidelijk, dat men van geen enkele signaalbeweging kan zeggen dat zij zeker géén oversprong-, intentie-, of ontwakende beweging is, vóórdat men deze beweging grondig vergelijkend bij een aantal soorten onderzocht heeft. Elke signaalbeweging kàn uit een dezer bronnen ontstaan zijn. Dit betekent natuurlijk ook weer niet, dat er geen andere bronnen van signaalbewegingen zijn; het veelvuldig voorkomende zich-groter-maken bijv., dat bij dreigende of sexueel imponerende dieren zo dikwijls voorkomt, heeft wellicht een andere oorsprong, en zo is er ongetwijfeld nog veel dat ons onbekend is. Het is niet nodig, hierop | |
[pagina 155]
| |
dieper in te gaan; de bedoeling was slechts, te demonstreren, hoe men door vergelijkende studie tot zekere gedachten kan komen over de wijze waarop signaalbewegingen zich ontwikkeld kunnen hebben. De oogst was mager wat conclusies betreft, maar dat ligt daaraan, dat het terrein nog niet heel intensief ontgonnen is. Maar, om bij deze beeldspraak te blijven, de verkaveling is toch al begonnen, en enkele wegen begeven zich al een eindweegs in het nieuwe land. Het gaat er dus maar om, ze te verlengen en te verbeteren, en tot een systeem samen te voegen. |