
Inleiding tot de diersociologie
(1946)–Niko Tinbergen–
[pagina 108]
| |
IV. De sociale structuur als geheel beschouwdTotnutoe hebben we ons, wat de samenwerking tussen individuen betreft, uitsluitend met de studie der eenvoudige relaties tussen twee dieren beziggehouden, met relaties die gebaseerd zijn op één signaalhandeling en één daardoor opgewekte reactie. Deze zeer eenvoudige relaties zijn echter niet meer dan de bouwstenen ener gemeenschap. Om de oorzakelijke structuur van een gemeenschap te leren begrijpen, om te zien hoe het uit bouwstenen opgetrokken geheel ‘in elkaar zit’, moeten wij niet alleen de bouwstenen zelf, maar ook hun samenvoeging tot een architectonisch geheel bestuderen. Wij moeten dus nu gaan onderzoeken, hoe uit de signaal-reactie relaties een gemeenschap opgebouwd kan zijn. Hiermee betreden wij nog weinig ontgonnen gebied. Moest ik bij de behandeling der eenvoudige relaties al op het fragmentarische karakter van het beschikbare feitenmateriaal wijzen, op het terrein dat wij nu betreden hangt onze kennis nog veel minder samen. Daardoor krijgt dit hoofdstuk meer nog dan het vorige het karakter van gedachtengangen, hier en daar door feiten geïllustreerd.
Wanneer wij ons niet, zoals in het vorige hoofdstuk, met opzet beperken tot het beschouwen van de eenvoudige relaties, dan komen wij als vanzelf ook met de meer ingewikkelde in aanraking. In het vorige hoofdstuk was de kernvraag, hoe een bepaalde reactie, bv. het eileggen van een stekelbaarswijfje veroorzaakt wordt. Wanneer wij ons in plaats daarvan afvragen, hoe een bepaald eindeffect tot stand komt, dus in dit geval, hoe het komt, dat er in het stekelbaarsnest bevruchte eieren komen, dan worden wij ter beantwoording van die vraag gedwongen, niet na de ontdekking van één relatie te stoppen, maar | |
[pagina 109]
| |
het onderzoek voort te zetten, tot alle relaties en hun samenvoeging ons duidelijk geworden zijn. In den regel zal men dan ontdekken, dat men met een aaneenschakeling van eenvoudige relaties te doen heeft, met een successie van signaal- en antwoordhandelingen. Bizonder duidelijk is dit bij het paargedrag van dieren. Wij blijven bij ons voorbeeld van de stekelbaars. De voorbereidingen voor de voortplanting treffen mannetje en wijfje afzonderlijk. Het mannetje kiest zich een territorium en bouwt er een nest, waarna hij bereid is, een wijfje te ontvangen. Het wijfje wordt drachtig en in haar rijpt de neiging, op een zigzaggende roodkaak toe te zwemmen. Hun gemeenschap begint bij de eerste reactie van het mannetje op het wijfje, de zigzagdans. Zoals wij gezien hebben, is door experimenten komen vast te staan, dat de zigzag-dans een reactie op een niet-rode stekelbaars met een dik achterlijf is. Als het mannetje nu zigzagt, gaat het wijfje ‘baltsen’: het zwemt in een bepaalde in hoofdstuk III beschreven houding op het mannetje toe. Dit baltsen is, zoals weer door proeven uitgemaakt is, een reactie op het zigzaggen. Komt het wijfje baltsend op het zigzaggende mannetje toe, dan keert hij om en zwemt snel naar het nest. Dit gedrag is een reactie op het toezwemmen en het volgen van het wijfje. Dit blijft, hiertoe Fig. 69. Het roodkaakje wijst een wijfje de opening van het nest (Ter Pelkwijk).
| |
[pagina 110]
| |
 Fig. 70. Het roodkaakje wijst een wijfje de opening van het nest (van boven gezien) (Ter Pelkwijk).
geprikkeld door het wegzwemmen van het mannetje, volgen. Ook dit volgen kan door op de juiste wijze bewogen marionetten opgewekt worden. Komt het wijfje zodoende dicht bij het nest, dan is haar aanwezigheid daar er de oorzaak van, dat het mannetje de kop in de nestopening legt en dan, de rug naar haar toegekeerd, plat op zijn zij gaat liggen, ondertussen met de snuit in de nestopening stotend (fig. 69 en 70). Dit prikkelt het wijfje er toe, in het nest naar binnen te kruipen, en als zij binnen is, prikkelt zij het mannetje tot ‘sidderen’. Dit sidderen wekt, zoals we gezien hebben, het eileggen op, en de vers gelegde eieren stimuleren het mannetje tot bevruchten. Als het vrouwtje haar eieren gelegd heeft, zwemt ze weg en de betrekkingen tussen de twee zijn verbroken; de gemeenschap is weer uiteengevallen in twee individuen. Wij zien dus dat het eindresultaat bereikt wordt door een lange keten van reacties, waarbij afwisselend het mannetje op een, signaal van het wijfje reageert en omgekeerd. De hele keten ziet men op blz. 111. Elke pijl tussen twee handelingen betekent niet alleen dat de handelingen op elkaar volgen, maar ook dat de een de ander opwekt. Elke antwoordhandeling van een der | |
[pagina 111]
| |
 dieren is tegelijk het signaal voor de ander, om hierop weer een nieuwe handeling zijnerzijds te laten volgen. Laat een der partners dan ook na, op een signaal van zijn tegenspeler te reageren, dan hapert hiermee niet alleen zijn eigen gedrag, maar onmiddellijk daarna ook dat van zijn tegenspeler. Ongetwijfeld wordt de paring bij vele dieren op soortgelijke wijze voorbereid. Het aantal opeenvolgende signalen kan groter of kleiner zijn, maar steeds heeft de samenwerking het karakter van een kaatsspel. Een complicatie vindt men bij de gevallen, waar de partners elkaar niet om beurten, maar tegelijk beïnvloeden, zoals bijv. het geval is bij het kopwippen van de zilvermeeuwen, het verenpoetsen van de kluten en het kopindopen van de ganzen. Bij deze dieren herhalen beide partners eenzelfde signaalbeweging een groot aantal malen; zij verwachten aanvankelijk van de ander niet een nieuwe signaalbeweging maar slechts steeds een nieuwe herhaling. Beide worden langzamerhand door de langdurige summatie der prikkeling in de stemming gebracht waardoor zij in staat zijn met de volgende signaalbeweging te antwoorden. De rhythmen der zich steeds herhalende signaalbewegingen van de twee partners zijn hier onafhankelijk van elkaar; elk zet zijn kopwippen, resp. verenpoetsen resp. kopindopen zo lang voort, tot hij door zijn maat in de volgende phase gebracht wordt. Willen wij ook hier de vergelijking met een kaatsspel volhouden, dan moeten wij toch eerder denken aan een kaatsspel met meer dan één bal. | |
[pagina 112]
| |
Het gevaar van een dergelijke paringsinleiding is natuurlijk, dat beide partners niet tegelijk bereid tot de paring zijn. Hoe dit bezwaar ondervangen wordt, is niet bekend.
Reeds herhaaldelijk hebben wij geconstateerd, dat vergelijking van verschillende diersoorten met elkaar aan het licht brengt, dat er verscheidenheid is, dat elke diersoort zijn eigen manier heeft, om een bepaald resultaat te bereiken. Ook hier weer dwingt de vergelijking van deze twee wijzen van paring ons tot de conclusie, dat een bepaald effect, de paring, bij verschillende diersoorten op heel verschillende wijze bereikt wordt. Vele wegen leiden naar Rome. Deze conclusie geldt in de biologie zeer algemeen, niet alleen wat het gedrag der dieren betreft, maar in het algemeen voor alle levensprocessen. Wanneer men de oorzakelijke structuur van tot een bepaald resultaat leidende levensprocessen vergelijkenderwijze, dus bij vele verschillende diersoorten, bestudeert, kan men wel als motto kiezen: ‘het kan zó, maar het kan óók anders’. In de vormleer zowel als in de functieleer wordt dit al lang beseft. Ik geef enkele voorbeelden. Een tot goed vormenzien capabel oog bijv. wordt bij verschillende dieren op geheel verschillende manier tot ontwikkeling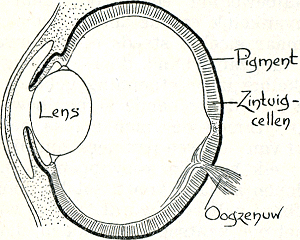 Fig. 71. Lensoog van een zoogdier (Kühn).
gebracht. Ons type, het op een camera gelijkende lensoog (fig. 71), komt bij vrijwel alle gewervelden en bij enkele groepen van ongewervelden voor. Bij een groep van inktvisschen komt een dergelijk type oog voor, dat geen lens heeft, maar gebouwd is volgens het principe van de ‘speldeprikcamera’: een kleine | |
[pagina 113]
| |
 Fig. 72. Het ‘speldeprikcamera-oog’ van Nautilus, een inktvis (Kühn).
opening heeft hetzelfde effect als een lens (fig. 72). Achter in het oog zit een ‘netvlies’, d.w.z. een vlies van lichtgevoelige cellen, waarop het door de lens resp. vooropening ontworpen beeld geprojecteerd wordt. Doordat dit beeld uit lichtere en donkerder partijen bestaat, worden sommige cellen sterk, andere minder sterk, andere, vrijwel of in het geheel niet geprikkeld. De totale prikkeling is dus a.h.w. een patroon van donkerder en lichtere punten, vergelijkbaar met een fijne zinkografieGa naar eind89. Bij geleedpotigen (kreeften, insecten) komt een heel ander oogtype voor: het facetoog (fig. 73). Hierin staan vele smalle, iets kegelvormige buisjes naast elkaar, door pigmentwanden optisch van elkaar geïsoleerd. Elk buisje is iets anders gericht dan zijn buurbuisje, en elk ontvangt dus licht van een ander deel van het gezichtsveld dan zijn buurbuisjes. Samen bestrijken zij een groot gezichtsveld. In elk buisje zitten lichtgevoelige cellen. Afhankelijk van de verdeling van donkere en lichte partijen in het gezichtsveld krijgen de verschillende buisjes een verschillende  Fig. 73. Facetoog van een insect (Kühn).
| |
[pagina 114]
| |
hoeveelheid licht. De lichtgevoelige cellen van elk buisje vormen hier nu één punt van de ‘zinkografie’, en zo ontstaat ook hier een projectie van de buitenwereld. Dit beeld is, in tegenstelling met het beeld in een lensoog, rechtopstaand, niet omgekeerd. Vervolgt men de structuur der ogen tot in détails, dan blijkt telkens weer, dat ook binnen één type verschillen bestaan, m.a.w. dat bepaalde effecten bij de ene soort zus, bij de andere zó bereikt worden. De instelling van de lens bijv., nodig om het beeld ook bij veranderende voorwerpsafstand steeds scherp te houden, kan òf door verschuiving van de lens, òf door vormverandering van de lens tot stand komen. Ook de verschuiving gaat niet bij alle dieren op dezelfde wijze. Vele vissen bijv. hebben de lens in rust ver van het netvlies en trekken hem achteruit wanneer zij een verwijderd voorwerp willen fixeren. Vele salamanders dragen de lens in rust dichter bij het netvlies en trekken hem naar voren wanneer zij iets van nabij willen bezien. Deze verplaatsingen geschieden door spieren, die aan de lens zelf bevestigd zijn; er zijn echter ook dieren waarbij drukverhoging in de oogbol de lens naar voren kan schuiven. En zo zijn er nog heel wat meer onderlinge verschillen. Een ander voorbeeld: de meeste vogels hebben aan hun poten drie naar voren en één naar achteren gerichte teen. Sommige echter hebben er twee naar voren en twee naar achteren gericht. Bij de spechten is het de buitenste der drie voortenen, die naar achteren gedraaid is, bij de in tropische oerwouden levende Trogonidae is juist de binnenste teen tot achterteen gewordenGa naar eind90. Bij de insecten zijn de sprieten dragers van de reukzin en van een fijn ontwikkelde tastzin. In verband hiermee zijn zij dikwijls lang en beweeglijk en met zintuigjes bezaaid. Ook bij de orde der kevers is dit zo. Bij de spinnende watertor echter, de bekende nestbouwende kever onzer sloten, zijn de sprieten als hulporganen bij de ademhaling in gebruikGa naar eind91. De functie van reuk- en tastorgaan wordt hier door de lip- en kaaktasters vervuld. Ook hier dus weer: het kan zó, maar het kan ook anders. | |
[pagina 115]
| |
Weer een ander aantrekkelijk voorbeeld levert de vergelijking van de bouw van de tong bij honingzuigende vogelsGa naar eind92. Deze gewoonte komt niet alleen bij kolibri's voor, maar ook bij sommige zangvogels. Bij beide groepen is de tong een lange holle buis, die zich aan de spits in twee of ook wel vier buisjes vertakt. Bij de honingzuigende zangvogels wordt de buis vrijwel geheel door verhoornde huidplooien gevormd; het tongbeen reikt niet verder dan tot de basis van de buis; bij de kolibri's zet het tongbeen zich tot in de toppen der buisjes voort en wordt de buis door zijdelings omkrullende hoornplooien gesloten. Bij de kolibri's dus tongspitsen van kraakbeen, dat de dunne lamellen steunt, bij de zangvogels tongspitsen van stevig hoorn, dat geen steun van node heeft.
Op sociologisch gebied is onze kennis van dit verschijnsel der convergente structuren nog zeer gebrekkig. Toch kan ik in aansluiting op wat ik hierboven over de verschillende copulatiemethoden schreef, enkele voorbeelden geven; mogelijk wekken zij tot verder onderzoek in deze richting op. Het is bekend, en ik heb nog met enkele voorbeelden gestaafd, dat elk vogelpaar in de regel zijn eigen jongen opvoedt. Bij zilvermeeuwen en sterns komt de hiervoor nodige binding aan de eigen jongen inderdaad door persoonlijk herkennen tot stand; de oude meeuw onderscheidt het eigen jong van vreemde jongen ook als deze laatste tussen zijn eigen jongen op zijn eigen territorium rondlopen. Een merel voedt óók uitsluitend zijn eigen jongen; hier is echter geen sprake van een persoonlijk herkennen. Hier komt de beperking der ouderlijke gedragingen tot stand door een sterke binding aan de plaats van het nest. Men kan de jongen door even oude jongen van andere merelparen vervangen, zonder dat de ouders blijk geven, iets van de verandering te merken. Wanneer een mensenkind naar zijn moeder loopt omdat zij tegen hem zei: ‘kom eens hier’, zien wij een sociolo- | |
[pagina 116]
| |
gische structuur in werking die hetzelfde effect bereikt als in een meeuwenfamilie de ‘au-roep’ van een der ouders, die de jongen naderbij lokt. Een mensenkind moet de betekenis der woorden leren, de jonge zilvermeeuw heeft een aangeboren gevoeligheid voor het signaal van zijn soort. Hetzelfde effect wordt dus weer op verschillende manieren bereikt. Bij de beschouwing van de rol der zintuigen bij sociologische connecties zijn wij eveneens op dit verschijnsel gestuit. Aanlokking van grote afstand gebeurt immers bij de dagpauwoog door middel van geurprikkels, bij sprinkhanen en zangvogels door geluidsprikkels, bij de heivlinder en bijv. ook de kamervlieg door gezichtsprikkels. Of men dus de gezichtszintuigen, het omklemmen van een tak door een vogelpoot, het aftasten van de geur der omgeving bij insecten, het honingzuigen bij vogels, het naar-zich-toe-roepen bij mensen en dieren beschouwt, steeds komt men tot de conclusie dat het zó kan, maar ook anders. In verband met het probleem dat ons in dit hoofdstuk bezighoudt, is het nu vooral interessant, om de oorzakelijke structuur van grotere sociologische complexen onderling te vergelijken. De meest bewuste poging in die richting is door Lorenz gedaan, die de paarvorming als geheel tot onderwerp van vergelijking maakte40). Hij kwam tot de conclusie, dat er betrekkelijk weinig verschillende manieren bestaan, waarop dieren in staat zijn de sexe van een hen ontmoetende soortgenoot te herkennen, en, wat voor ons hier het belangrijkste is, dat geen dier methoden karakteristiek is voor een systematisch bepaalde diergroep, doch dat integendeel alle drie methoden bij zeer verschillende diergroepen in zwang zijn. Zo vindt men drie typen, die bij vogels bekend zijn, ook bij vissen; één type is nu zelfs bekend van vogels, hagedissen, vissen en inktvissen. Bij de in hoofdstuk III genoemde leguaan Sceloporus undulatus dient de blauwe onderzijde in de eerste plaats er toe, te verhinderen dat een mannetje met een ander manne- | |
[pagina 117]
| |
tje, in plaats van met een wijfje, paart. De mannetjes dezer soort hebben namelijk de neiging, alle soortgenoten met een copulatiepoging te bejegenen, tenzij deze het typisch mannelijke signaal tonen. Een dergelijke wijze van paarvormen treft men ook bij sommige vogels aan, bv. bij de muskuseend, en ook bij de inktvis Sepia officinalis, onze gewone ‘zeekat’. Waarschijnlijk komt zij ook bij sommige vissen voor. Heel anders gaat de paarvorming bij sommige labyrinthvissen toe, bijv. bij de Indische kempvis. Hier willen de mannetjes in eerste instantie tegen elk dier vechten, en tot een paring komt het slechts met een soortgenoot, die de voor een wijfje karakteristieke signalen demonstreert. Deze signalen zijn dikwijls gelijk aan die waarmee het dier aangeeft dat het zich sociaal aan de ander onderwerpt; het wijfje moet zich dus in de sociale rangorde onder het mannetje willen plaatsen, anders loopt de paarvorming mis, doordat het mannetje tot vechten geprikkeld wordt. Ook deze wijze van paarvorming komt bij vele vogels voor, bv. bij kauwtjes, raven en duiven. Bij sommige Cichliden komt de paarvorming weer anders tot stand. Het wijfje draagt hier hetzelfde kleed als het mannetje en imponeert tegenover hem op dezelfde wijze als hij tegenover haar. Voor een goede gang van zaken is hier niet nodig dat het wijfje zich de sociaal lagerstaande toont, integendeel, àls zij dat zou doen, zou het mannetje haar niet als wijfje accepteren. Zij moet terug imponeren, ‘pronken’ en pas na lang wederzijds pronken komen de partners langzamerhand in de stemming waarin zij het, uiteindelijk tot de paring leidende, vraag- en antwoordspel kunnen inzetten. Deze paring zonder rangorde en met wederzijds imponeren kent men onder de vogels bijv. bij reigers. Deze vergelijking is, het moet toegegeven worden, nog maar heel schetsmatig, om de eenvoudige reden, dat voor preciesere en bredere vergelijking de nodige feiten nog ontbreken. Het moge echter duidelijk zijn, dat de paarvorming op meer dan één wijze tot stand kan komen, en | |
[pagina 118]
| |
verder, dat bepaalde typen van paarvorming zich bij verschillende, onderling niet bepaald nauw verwante soorten ontwikkeld hebben. Gelijkheid in paarvormingstype wijst geenszins op verwantschap, maar alleen op, in dit opzicht, gelijkgerichte of beter convergente ontwikkeling. Nog één ander voorbeeld van convergentie in grotere sociologische complexen moet ik hier noemen: de paralleliteit in de arbeidsverdeling in mieren- en termietenstaten. Hoewel termieten en mieren met elkaar helemaal niet nauw verwant zijn (termieten zijn verwant met kakkerlakken, mieren met de in vele opzichten hoger ontwikkelde bijen en wespen), heeft het sociale leven dat beide groepen leiden, dusdanig zijn stempel op hen gedrukt, dat de organisatie hunner staten en de er heersende arbeidsverdeling een verbazende gelijkenis vertonen. Evenals bij de mieren zijn er bij de termieten speciale individuen die voor de voortplanting zorgen: bij de mieren heeft elk volk een koningin, bij de termieten (waar alle kasten uit mannetjes en wijfjes bestaan) een ‘koningspaar’. Evenals bij de mieren vindt men bij de termieten ‘werkers’ en ‘soldaten’. Er zijn mieren die een soort landbouw beoefenen in de vorm van het kweken van schimmels; er zijn eveneens, schimmelkwekende termieten. Zowel de schimmelkwekende mieren als hun collega's onder de termieten knippen met hun kaken stukjes uit grassprieten en andere bladeren en gebruiken deze als mest voor hun schimmels. De ‘soldaten’ zijn, zoals op bladzijde 101 al beschreven werd, zowel bij mieren als bij termieten uitgerust met grote koppen en zware kaken, en hebben dikwijls een speciale taak bij de verdediging van de staat. Uit deze voorbeelden blijkt wel, dat ook in de sociologie geldt: een bepaald effect kan steeds op meer dan een wijze bereikt worden, ja men kan wel zeggen, dat geen twee diersoorten hetzelfde effect op precies dezelfde wijze bereiken. Dit leidt naar een voor de hand liggende conclusie, n.l. dat zulk een effect blijkbaar iets heel noodzakelijks is, ‘dat het om het effect te doen is’. In vele gevallen neemt men dat voetstoots aan: niemand twijfelt | |
[pagina 119]
| |
er aan, dat een oog een hoogst noodzakelijk orgaan is, of dat het van groot belang voor een kolibri is, om een instrument te bezitten waarmee hij kan honingzuigen. Het behoud van het individu is immers van het goede functionneren van deze organen afhankelijk. De vraag rijst nu: is het behoud van het individu óók van het goede functionneren der sociologische ‘uitrusting’ afhankelijk? Is ook in de sociologie het ‘nut’ van de ingewikkelde mechanismen die alle naar bepaalde effecten streven, evident? Het blijkt al bij oppervlakkige beschouwing, van de in de vorige hoofdstukken beschreven voorbeelden dat van voordeel voor het individu dikwijls geen sprake is. Wanneer een sociale vogel een koloniegenoot voor gevaar waarschuwt, is dit voor hem zelf van geen nut, integendeel, hij vestigt hoogstens de aandacht van een roofvijand op zichzelf. Wanneer een kievitvader zijn jong uit de nabijheid van een kat wegjaagt, begeeft hij zich in direct gevaar. Het is duidelijk, dat vele sociale signalen voor de actor zelf eerder gevaarlijk dan nuttig zijn. Zij zijn echter nuttig voor de grotere, sociale eenheid, voor de gemeenschap die er in betrokken is. Een alarmroep, een sociale aanval kan koloniegenoten het leven redden, een signaal om nu en hier eieren te leggen (stekelbaars) vergroot de kans op bevruchting en verdere verzorging der eieren zeer. De signalen mogen dus al niet het individu dienen, zij dienen stellig de gemeenschap. Die gemeenschap is in het ene geval groter dan in het andere. Nu eens gaat het om het paar, dan om het gezin, dikwijls ook om een grote groep van soortgenoten. Steeds dienen zij daarbij de soort, d.w.z. zij dragen bij tot zijn instandhouding. Eén categorie van betrekkingen, die ik in de vorige hoofdstukken ook al min of meer als een buitenbeentje behandeld heb, moet hier nu nog nader bezien worden: de gevechten, die, zoals gezegd, in het dierenrijk vrijwel uitsluitend aan de paarvorming gebonden zijn. Is er ook bij de paarvormingsgevechten, bij de verdediging van vrouwtje, territorium of andere voor de voortplanting noodzakelijke voorwerpen sprake van voordeel voor de soort? | |
[pagina 120]
| |
Men meent van wel, en de juistheid hiervan blijkt wel, wanneer men eens nagaat wat er zou gebeuren als deze vechtdrang er tijdens de paarvorming niet was. Het meest voorkomende geval is hier het verdedigen van een territorium. Wanneer een bepaald gebied in het voorjaar door zangvogelmannetjes bezet wordt, gaat dit geleidelijk in zijn werk. Eerst zijn er weinig mannetjes, elk met een groot territorium; dan vestigen zich er meer, die tijdens hevige gevechten stukjes van de rand van reeds bezette territoria afknabbelen, en zo zelf een territorium krijgen. Dat dit in den beginne nog kan, ligt daaraan dat de vechtdrang der territoriumbezitters bij een groot territorium gering isGa naar eind93. Omgekeerd geldt: hoe kleiner het territorium wordt, des te feller wordt het verdedigd. Tenslotte, als zich meer en meer vogels gevestigd hebben, worden de territoria zó klein en de vechtlust der eigenaars daardoor zó groot, dat geen nieuweling zich meer kan indringen. Er komen dus in dit terrein een bepaald, gelimiteerd aantal vogels te wonen, die er voor zorgen, dat het gebied niet dichter bevolkt wordt. Met andere woorden, de sexuele vechtdrang voorkomt overbevolking. Men heeft zich, in het geval der ‘territoriumvogels’ wel eens afgevraagd, in welk opzicht overbevolking schadelijk zou zijn, of in andere woorden, waarvoor het territorium dient. Dit is stellig van soort tot soort verschillend. Voor vele zangvogels waarborgt het territorium een bepaalde hoeveelheid voedsel dicht bij huis, zodat de ouden niet ver weg hoeven te vliegen om het te halen, en dus niet lang van het nest hoeven weg te blijven, wat vooral van belang is zolang de jongen nog klein zijn en geregeld gekoesterd moeten worden. Bij andere vogels (reigers, sterns, zeekoeten) levert het territorium geen voedsel, maar is het als nestplaats van waarde. Welke echter ook de schadelijke gevolgen van overbevolking in de afzonderlijke gevallen zijn, het zit in de term opgesloten dat overbevolking schadelijk is, en het is duidelijk dat de verdediging van een territorium van voordeel voor de soort als geheel is. Er zijn echter ook soorten, wier vechtdrang onafhan- | |
[pagina 121]
| |
kelijk van een territorium is en er op gericht is, rivalen uit de nabijheid van de geslachtspartner te houden. Men neemt ook hier aan, dat het vechten van belang voor de soort is, daar het promiscuiteit voorkomt. Het is in dit bestek niet mogelijk, hierop nader in te gaan, maar op één duidelijk, veel voorkomend geval wil ik toch wijzen. Bij soorten waar zowel de mannetjes tegen de mannetjes als de vrouwtjes tegen de vrouwtjes vechten, wordt hierdoor monogamie bevorderd, want zodra zich bij een paar een vreemde zou willen voegen, wordt hij door één van beide partners aangevallen. Monogamie is bij vele soorten van groot belang voor de broedzorg, nl. bij die soorten waar man en vrouw samen de jongen verzorgen. Bigamie zou daar bv. meebrengen, dat het mannetje zijn aandacht over twee broedsels zou moeten verdelen. Bij bigame mannetjes van normaliter monogame vogels heeft men inderdaad waarnemingen verricht die er op wijzen dat hun broedsels minder goed slagen dan die der monogame mannetjesGa naar eind94.
Minder zeker is men bij de beoordeling van de vraag of de gevechten die tot het vestigen van een rangorde leiden, van nut voor de soort zijn. Men zou zich kunnen voorstellen, dat door de rangorde een zekere rust binnen de troep bereikt wordt, die voorkomt dat allerlei voor de soort nuttige gedragingen door chaotisch gekrakeel verstoord worden. Men kan zich echter ook indenken dat het er voor de soort niets toe doet of er een rangorde heerst of wel anarchisme. Het woord anarchisme noopt echter al dadelijk tot het trekken van een vergelijking met de menselijke sociologie, en hier is men het er wel over eens, dat anarchisme onmogelijk van voordeel voor de gemeenschap kan zijn. De vaagheid van dit betoog toont echter al, hoe weinig men nog over de diersociologische zijde van deze kwestie oordelen kan.
Onze slotconclusie omtrent de kwestie van het nut moet, ondanks de laatst besproken onzekerheid, toch wel luiden, | |
[pagina 122]
| |
dat sociale betrekkingen van nut zijn voor het behoud van de soort. Men kan zelfs nog verder gaan. Er bestaan processen die hun nut niet voor een soort afwerpen, maar voor meer dan een soort. Het duidelijkste voorbeeld hiervan is de ‘fremddienliche Zweckmässigkeit’, die men in de plantengallen ontdekt heeftGa naar eind95. Vele plantengallen vormen niet zonder meer een wondweefsel dat de wondveroorzaker moet inkapselen en onschadelijk maken, maar zijn integendeel uitstekende beschermende tehuizen voor de wondverwekkers. Het vreemde daarbij is, dat de dienstbaarheid zo eenzijdig is; het is niet in te zien hoe de plant voordelen uit de galverwekker zou trekken. De enige conclusie die men kan trekken is, dat de gal een gemeenschap tussen twee soorten is, die om haarszelfs wille behouden blijft en daartoe door aanpassingen van de zijde van de plant gediend wordt. Het is zeer goed mogelijk, ja waarschijnlijk, dat dergelijke gemeenschappen van hogere orde, waarin soorten offers brengen tot heil van de grotere gemeenschap, zoals in sociale complexen de individuen offers brengen tot heil van de soort, en zoals in het individu organen offers brengen tot heil van het individu, veel meer voorkomen. Het gaat er maar om, de natuur vanuit dit gezichtspunt te bezien.
Deze beschouwingen leiden er als vanzelf toe, om het sociale complex als organisme, als individu van hogere orde te gaan beschouwenGa naar eind96. Of om omgekeerd datgene wat we gewoonlijk als individu beschouwen, als georganiseerde gemeenschap te bezien. Dat wil dus zeggen, dat wij zowel het individu als de kolonie als een vorm van samenwerking tussen elementen, organen, gaan beschouwen. Tot deze beschouwingswijze komt men eerst recht, wanneer men door vergelijkende studie ontdekt, dat er in het dierenrijk organismen voorkomen, waarvan men niet weet of men het individu of kolonie moet noemen. Er zijn nl. individuen die de eigenschappen van een kolonie vertonen, en ook kolonies die de eigenschappen van een individu vertonenGa naar eind97. Als kriterium nemen wij de | |
[pagina 123]
| |
in de naam ‘individu’ besloten eigenschap van ondeelbaarheid. Men neemt van een individu aan, dat de samenstellende delen slechts in hun verband met elkaar, dus als geheel kunnen leven. Tot op zekere hoogte is dat juist; een van ons lichaam afgesneden vinger bijv. kan niet zelfstandig verder leven. Er zijn echter bij de lagere dieren velerlei soorten waarvoor dit niet geldt. Een worm bijv. of een zoetwaterpoliep kan men in vele stukken snijden zonder die stukken daarmee te doden; elk ontwikkelt zich in korte tijd tot een nieuw, volledig individu. Zijn zulke ‘individuen’ dus niet eigenlijk ‘celstaten’? Dat wij ook cellen als individuen kunnen beschouwen, bewijzen ons immers de eencelligen. Een pantoffeldiertje is stellig niet minder een individu dan bijv. een zeester. Men kent kolonies van eencellige dieren, die zó duidelijk met ‘niet-sociale’ eencelligen verwant zijn, dat men niet aarzelt, hen kolonies en geen veelcellige dieren te noemen (fig. 74); maar dit wordt al heel wat problematischer, wanneer in zulk een kolonie een verdeling van arbeid bestaat tussen cellen die voor de voeding zorgen en cellen die voor de voortplanting zorgenGa naar eind98. Moet men de afzonderlijke cellen hier nog individuen noemen, of reproductieve resp. vegetatieve organen? 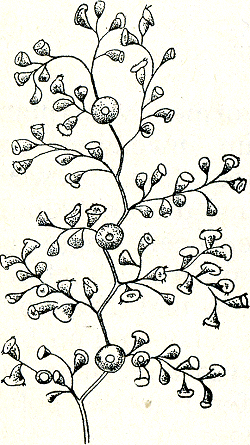 Fig. 74. Een protozoënkolonie.
Aan de andere kant kent men vele dierenstaten, waarvan de leden, elk een afzonderlijk veelcellig individu, geenszins een afzonderlijk bestaan kunnen leiden. Er bestaan bijv. kolonies van poliepachtige dieren, waarvan de afzonderlijke individuen met elkaar vergroeid zijn. Scheidt men één individu hiervan af, dan gaat dit te gronde. Men kent onder die kolonies zelfs vormen, de in zee levende Siphonophoren (fig. 75), waarvan de verschillende individuen onderling grote verschillen in bouw vertonen; sommige zijn in bouw en functie gespecialiseerd op het opnemen van | |
[pagina 124]
| |
 Fig. 75. Een Siphonophore (Koelliker).
voedsel voor de hele kolonie, andere vormen voortplantingscellen, weer andere vormen in hun inwendige een gasbel, die de hele kolonie drijvend moet houden, weer andere zijn gewapend met giftige wapens, die de kolonie tegen roofvijanden moet beschermen. Men heeft lange tijd in het onzekere verkeerd, of men hier te doen had met één individu met organen, òf met een kolonie met ‘kasten’. Slechts door vergelijkende studie, van deze soorten en hun verwanten, ontdekte men dat er zekere tussenvormen bestaan tussen dit type van sterkst gedifferentieerde poliepenkolonie en dat van de solitaire poliepenGa naar eind99. Ook andere dierenkolonies zijn, stellig in samenhang met de mate van arbeidsverdeling, min of meer ‘ondeel- | |
[pagina 125]
| |
baar’. Een bijenvolk bijv. kan niet willekeurig in delen gesplitst worden. Noch de koningin, nòch de werksters, nòch de darren kunnen alléén leven; bij het zwermen, de natuurlijke splitsing van de kolonie, delen dan ook alle kasten zich in twee groepen. En niet alleen bijen zijn tot individueel leven incapabel; ook andere sociale dieren waarvan men het op 't eerste gezicht niet verwachten zou, kunnen niet of slecht individueel leven. ‘Een eenzame chimpansee is geen chimpansee’, zegt de beste kenner dezer dieren, Wolfgang Koehler zelfs, waarmee hij bedoelt, dat een chimpansee zich pas volledig kan uitleven wanneer hij in de troep leeftGa naar eind100. En in hoe sterke mate geldt dit ook niet voor de mens! Verreweg de meesten van ons zouden, zelfs al zouden zij zich materieel kunnen helpen, in totale eenzaamheid min of meer kwijnen. Zowel bij de kolonies van sociale dieren als bij de kolonies van sociale cellen (om de veelcellige dieren maar eens zo te noemen) vinden wij dus gevallen van sterker en minder sterke samenvoeging, van slechtere en betere deelbaarheid. Wij komen dus tot de conclusie, dat er nòch tussen eencellig organisme en celstaat, nòch tussen veelcellig organisme en kolonie scherpe scheidingen bestaan. Wij kunnen zelfs verder gaan, en deze toestand dynamisch interpreteren, als resultaat van een ontwikkelingsgang, die er op gericht is, organismen tot organismen-complexen, tot ‘superorganismen’ te doen samengroeien. Deze ontwikkeling bestaat uit een samenwerking van individuen, hand in hand gaande met een verdeling van arbeid, met differentiatie dus. Hoe sterker de differentiatie, hoe nauwer de samenwerking, des te sterker krijgt de ‘kolonie’, de ‘staat’ of hoe men het samenvoegsel ook noemen wil, de eigenschap van ondeelbaarheid, van het superorganisme, van het individu. Zo heeft, naar men inderdaad op goede gronden kan aannemen, deze ontwikkelingsgang er toe geleid, na en uit eencelligen de veelcelligen, uit veelcelligen de kolonies en staten te doen ontstaan. En het is verleidelijk, deze ontwikkelingsgang ook in de menselijke sociologie te zien, en te zien hoe hij als 't ware voortgezet | |
[pagina 126]
| |
wordt; verscheidene moderne staten zijn uit een vereniging van kleine staatjes ontstaan, en zelfs de grote staten zijn onderling tot een groter geheel, althans tot enkele grotere eenheden verbonden. Zo ligt zoiets als de Volkenbond of de U.N.O., biologisch gezien, in de lijn der natuurlijke ontwikkeling. Deze beschouwingswijze is meer dan alleen-maar een aardige manier om een kolonie te zien. Zij geeft ons een methode aan, die wij bij het onderzoek van kolonies kunnen toepassen. Precies zoals we bij een individu de structuur en zijn werking onderzoeken, de afzonderlijke functies in hun werking trachten te begrijpen, zo kunnen we dat ook bij de superorganismen doen. Hoe wordt de kolonie beschermd, zoals huid en verdedigingsorganen het individu beschermen? Waar zijn de zintuigen van de kolonie, waar de voedingsorganen, waar de voortplantingsorganen? We kunnen dus de organen van individu en super-individu vergelijken, zoals men dat reeds lang doet met de organen van een individu en de ‘organellen’ van een eencellige. Men vindt dan inderdaad een goede weg tot analyse der sociale structuur, en al onderzoekende stuit men op merkwaardige parallellen.
Beschouwen wij eerst de verdeling van arbeid nader. Zoals het individu er verschillende organen op na houdt, die uit verschillende celsoorten bestaan, die binnen het orgaan op een eigen manier samenwerken, zo heeft de individuenkolonie of -‘staat’ zijn ‘organen’, die uit speciale individuen bestaan, die dikwijls op speciale wijze samenwerken. Elk dier heeft bijv. zijn organen voor het opnemen van het voedsel, en ieder weet hoe hoogst ingewikkeld de structuur en werking dezer organen zijn kunnen, men denke bv. aan de schoolboekjesvoorbeelden der insectenmonddelen. Een orgaan voor voedselzoeken vindt men ook in de bijenstaat. Hier zijn het een deel van de werksters, en wel, zoals gezegd, een bepaalde leeftijdsklasse, die uitvliegen en honing en stuifmeel brengen. Bij mieren en termieten | |
[pagina 127]
| |
is er eveneens een kaste van voedselzoekers, en hier zijn deze dieren zelfs in bouw verschillend, niet alleen van de voortplantingsdieren, maar ook van de verdedigingsdieren, de ‘soldaten’. Bij de Siphonophorenkolonie zijn er, zoals gezegd, ook bepaalde individuen, die uitsluitend voedsel opnemen en die in bouw van de andere ‘organen’ der kolonie verschillen. Op dergelijke wijze bestaat er een parallel tussen de geslachtsorganen der individuen en die van een kolonie. Darren en koninginnen van de bijenstaat, koning en koningin der termieten zijn de ‘geslachtsorganen’ der staten. Ook op het gebied der zintuigen kan men de parallel trekken: de zoekerbijen, de pioniers die nieuwe voedselbronnen ontdekken, zijn niet anders dan de naar voren geschoven zintuigen van de kolonie, die in dienst van het geheel staan. Zoals een slak zijn voelhorens uitsteekt, waarop zijn ogen en zijn tastzintuigen zetelen, en zich dus door middel van enkele lichaamsdelen een beeld van zijn verdere omgeving kan vormen, zo stuurt een bijenstaat zijn zoekers uit om de verdere omgeving te onderzoeken. Zoals het zenuwstelsel van de slak de boodschap der zintuigen doorgeeft naar de effectoren, de spieren, die voor het binnenbrengen van het gevonden voedsel zorgdragen, zo zorgt de signalentaal der dansende pioniersbijen er voor, dat de effectoren van de staat, het leger der honinghaalsters, in beweging komt en het voedsel binnenbrengt. Zoals de huid van een dier zijn lichaam tegen schadelijke invloeden beschermt, zo beschermt de wachterbij het volk tegen vreemde indringers. Zoals bepaalde netelcellen in de huid van een poliep of kwal roofvijanden doen terugdeinzen, zo doen de neteldieren der Siphonophoren, de soldaten der termieten dat. Afvalstoffen worden door nieren en hun hulporganen uit het lichaam verwijderd; een bepaalde leeftijdsklasse onder de bijen zorgt voor de verwijdering van de afvalstoffen uit de korf. En zo zou men verder kunnen gaan; de verdeling van arbeid in de staat volgt dezelfde principes als de verdeling van arbeid in het individu. | |
[pagina 128]
| |
Even leerzaam is het, de samenwerking tussen de organen bij een celstaat en een individuenstaat te vergelijken. Samenwerking tussen de organen van een celstaat is alleen mogelijk doordat het ene orgaan het andere beïnvloedt. Men kent twee wijzen van beïnvloeding: die door middel van het zenuwstelsel en die door middel van stoffen die door het bloed van het ene orgaan naar het andere gevoerd worden, zgn. hormonen. De beïnvloeding door zenuwimpulsen voltrekt zich in zeer korte tijd, en een zintuig kan dan ook de samentrekking van spieren bewerken in een onderdeel van een seconde. De hiermee vergelijkbare functie in het superorganisme wordt verricht door de signalentaal, waarmee het ene individu het andere beïnvloedt. Het sperren van de jonge zangvogels roept een onmiddellijke reactie in de oudervogel op; het opvliegen van één taling in een groep doet de hele troep opvliegen. De beïnvloeding door hormonen gaat langzamer, ten eerste doordat het transport er van met niet groter snelheid dan die van de bloedstroom geschiedt, ten tweede omdat de reactie meestal niet uit ogenblikkelijke spiersamentrekkingen bestaat maar uit het in werking treden van een klier of uit zich langzaam voltrekkende groeiprocessen. Er zijn processen in het superorganisme die enigszins hiermee te vergelijken zijn. Als voorbeeld noem ik de beinvloeding van de eiontwikkeling bij duiven door de baltsceremoniën van de doffers. Wanneer men een pronkende doffer in de buurt van een wijfje brengt, maar hun verhindert haar te bevruchten, dan kan alleen de aanblik van de doffer haar al tot ontwikkeling van de eieren, ja tot eileggen brengen. Een gevolg van deze omstandigheid is het ook, dat, wanneer men twee wijfjes bij elkaar opsluit - waarbij steeds één wijfje zich als mannetje gaat gedragen en de ander het hof gaat maken - het bij zulk een wijfjespaar tot nestelen en eileggen komt, hoewel er toch van een bevruchting nooit sprake geweest is. Hieruit blijkt, dat ook bij de normale paarvorming bij duiven lang- | |
[pagina 129]
| |
zame invloeden van het ene dier op het andere werken. Ook in het superorganisme - hier: het paar - kent men dus een dergelijke invloed van het ene orgaan (de doffer) op de groeiprocessen in het andere (de duif). Ook hier gaat de beïnvloeding via signalen.
Een merkwaardige parallel bestaat er tenslotte ook tusssen de wijze waarop in celstaat en in individuenstaat de differentiatie der organen ontstaat. De studie van de ontwikkeling van de lens van het oog bij Gewervelden bijv. heeft aan het licht gebracht dat de vóór het zich ontwikkelde oog liggende huid zich onder invloed van stoffen, die van het groeiende oog uitgaan, tot een lens ontwikkelt. Men heeft deze beïnvloeding ontdekt door proeven waarvan de volgende er één is. Brengt men operatief de oogaanleg op een verkeerde plaats van het lichaam, bijv. onder een stuk huid van de rug, dan ontwikkelt de huid juist hier zich tot een lens, terwijl dan op de plaats vanwaar men de oogbeker wegnam en waar zich normaliter de lens vormt, geen lens tot ontwikkeling komtGa naar eind101. Een dergelijke chemische beïnvloeding van de groei van een orgaan kent men ook in het superorganisme. Bij de honingbij bijv. zijn de eieren, waaruit later koninginnen zullen komen, in erfelijke aanleg gemiddeld precies gelijk aan die waaruit de werksters moeten komen. Door (kwantitatief èn kwalitief) verschillend voedsel echter ontwikkelen sommige eieren zich tot de geslachtsdieren, andere tot de, wat de geslachtsfuncties betreft gedegenereerde, in andere opzichten echter gespecialiseerde, werksters. Bij de termieten neemt men zelfs op goede gronden aan, dat ook de verschillen tussen de kasten der werkdieren onder de chemische invloed van verschillende groeiregelende stoffen tot stand komen.
Wanneer in de celstaat één orgaan of een deel er van uitvalt, neemt dikwijls een ander deel zijn functie geheel of gedeeltelijk over, ook al oefent het in het normale individu die functie niet uit. Bekend is bijv. dat er tussen de | |
[pagina 130]
| |
verschillende delen van de schors der grote hersenen een verdeling van arbeid bestaat; men kan ‘velden’ met bepaalde functies aanwijzen. De ervaring heeft nu geleerd, dat bij beschadiging van sommige zulke velden de functies, die eerst gestoord zijn, langzamerhand weer normaal vervuld worden, en wel doordat andere, onbeschadigd gebleven delen die functies overgenomen hebbenGa naar eind102. Dergelijke verschijnselen vertoont het superorganisme. Bij vele roofvogels bijv. heerst tijdens het broedseizoen verdeling van arbeid tussen man en vrouw. Het mannetje jaagt en brengt het voedsel aan het vrouwtje, dit verdeelt het voor de jongen en koestert en bewaakt de jongen ook. Deze arbeidsverdeling is zó rigoureus, dat, als het wijfje tijdens deze phase om het leven komt, het broedsel in de regel verloren gaat. In enkele gevallen heeft men echter waargenomen, dat het mannetje na enige tijd de taak van het verongelukte wijfje overnam en de jongen toch ging voerenGa naar eind103. In dit verband zijn ook de eigenlijke regeneratieverschijnselen interessant. Bij vele dieren kunnen verloren gegane lichaamsdelen weer aangroeien. Verliest een salamander bijv. een vinger, of zelfs een poot, dan groeit dit lichaamsdeel na enige tijd weer normaal aan. Dergelijke regeneratieverschijnselen kent men ook in het super-organisme. Een heel boeiend voorbeeld hiervan levert hiervan de bijenstaat. Normaliter bestaat er, zoals op bladzijde 13 beschreven, tussen de werksters een, op leeftijdsverschil berustende arbeidsverdeling, waarbij de verschillende sociale functies elk door een bepaalde scherp begrensde leeftijdsklasse vervuld worden. Schakelt men nu kunstmatig een of meer klassen uit (neemt men dus één der organen van het superorganisme weg) dan treden regeneratieverschijnselen op, die de storing weer ongedaan maken en daardoor het geheel, de staat, van een wisse ondergang reddenGa naar eind104. Zo kunnen bijv. bij het uitvallen der honing- en stuifmeelzoeksters, gewoonlijk dieren van twintig dagen en ouder, jonge bijen van amper zes dagen oud, die gewoonlijk binnen werkzaam zijn en de larven | |
[pagina 131]
| |
voeren, zich tot honinghaalsters opwerpen. Neemt men alle met de bouw der raten bezige bijen weg, die alle omstreeks achttien tot twintig dagen oud zijn, dan worden oudere bijen, die al tot honingzoeksters waren geworden (wat met degeneratie der was afscheidende klieren gepaard gaat) weer tot bouwsters; hun wasklieren regenereren en zij beginnen weer zelf te bouwen. Hoe deze regeneratie tot stand komt, welke prikkels de dieren tot afwijking van hun normale groei en gedrag brengen, is nog niet bekend. Juist als bij de regeneratieverschijnselen in de celstaat openen deze ontdekkingen ons een nieuw veld van onderzoek; in beide gevallen dringt zich de vraag naar de veroorzaking dezer, voor het geheel zo nuttige, processen op. Dat men in het geval van het superorganisme evenmin van ‘inzicht’ der organen in het door de regeneratie bereikte doel kan spreken als men dat bij de organen van de celstaat doet, behoeft, na het in de vorige hoofdstukken aangetoonde, wel geen betoog.
Deze vergelijkingen zijn met opzet zeer schematisch gehouden, daar het er niet om te doen was, feitelijke resultaten van een analyse te geven, maar om te tonen, hoe men, door het wijzen op de parallellen tussen organisme en superorganisme, in de sociologie tot dezelfde wijze van problemen stellen komt als in de physiologie van de cellenstaat. In de grond heeft men èn bij de studie van het eencellige wezen, èn bij de studie van de cellenstaat èn bij de studie van de individuenstaat met dezelfde problemen te maken: differentiatie (verdeling van arbeid)-tussen kern en plasma of tussen de organellen in het eencellige organisme; tussen de organen in het veelcellige organisme; tussen de kasten in de individuenstaat, het superorganisme - en samenwerking tussen deze differente onderdelen. Hoe ingewikkelder het organisme, hoe moeilijker deze kernproblemen te zien zijn. Bij het eencellig wezen liggen zij zeer voor de hand; bij het veelcellige dier reeds minder en bij het superorganisme kost het moeite, behalve de bomen ook het bos te zien; om niet alleen de individuen, | |
[pagina 132]
| |
maar ook het geheel te zien. Juist hier is het moeilijk, om tot nuchter, objectief onderzoek te komen en dat is de reden waarom ik hier juist veel ruimte besteed heb aan het trekken van parallellen tussen organisme en super-organisme. De vergelijking tussen organisme en gemeenschap, waardoor de gemeenschap als superorganisme herkend wordt, maakt dat de weg tot dit objectieve onderzoek zich plotseling als het ware duidelijk afgebakend aan ons voordoet. |
|