
Inleiding tot de diersociologie
(1946)–Niko Tinbergen–
[pagina 28]
| |
III. De analyse van de sociale structuurA. Inleiding.Bij de zo juist genoemde voorbeelden werd het al duidelijk, dat de gemeenschap bij dieren, juist als de menselijke samenleving, berust op een met-elkaar-in-verbinding-treden der individuen, op coördinatie en synchronisatie hunner gedragingen. En evenals bij de mensen wordt die samenwerking mogelijk gemaakt doordat de individuen elkaar bepaalde ‘mededelingen’ doen, elkaar ‘signalen’ geven, in zekere zin dus over een ‘taal’ beschikken. Ik plaats deze termen tussen aanhalingstekens om te doen uitkomen dat zij in de diersociologie een andere betekenis hebben dan in de mensensociologie - een kwestie waarop ik spoedig terug zal komen. Zulke ‘signalen’ zagen wij in het vorige hoofdstuk dan ook telkens in functie; herhaalde malen bleek immers dat A iets deed juist nadat B iets gedaan had, waarbij wij al dadelijk vermoedden, dat datgene dat B gedaan had als signaal voor A gewerkt had. Als bijv. de jonge zanglijster (B) zijn bek openspert, voert de oude (A) hem. Als de eene roodkaak (B) dreigend komt aanzwemmen, wijkt de ander (A) terug. Wil de diersocioloog inzicht in zulke gevallen van gemeenschapsleven krijgen, dan moet hij beginnen, dit proces, waarin speler en tegenspeler, actor en reactor, hun gedrag wederzijds op elkaar afstemmen, te analyseren. Hij moet weten, uit wat voor soort bewegingen het signaal bestaat, verder wat de actor drijft bij het geven van zijn signaal, en ook waardoor de reactor op de juiste, doeltreffende manier reageert, kortom hij moet de keten van processen uiteenrafelen, die begint bij het initiatief van de actor en eindigt bij de antwoordhandeling van de reactor. Niet alleen de samenwerking echter wil hij analyseren. Met samenwerking gaat steeds verdeling van functie hand | |
[pagina 29]
| |
in hand. Tussen de samenwerkende leden van een gemeenschap bestaat steeds verschil. Soms groot, zoals tussen mannetje en vrouwtje, soms kleiner, zoals tussen leiderhond en ondergeschikte hond, soms nog subtieler, zoals tussen de spreeuwen van een spreeuwenwolk, waarvan de één door zijn iets eerder zwenken de ander ten voorbeeld is. Nooit hebben twee samenwerkende individuen precies dezelfde functie. Deze differentiatie, deze arbeidsverdeling, te leren kennen is voor een goed begrip van een gemeenschap even belangrijk als het onderzoek van de samenwerking. | |
B. De samenwerking.Het programma omvat dus een onderzoek naar de samenwerking (waaronder ik óók de tegenwerking vat) en naar de arbeidsverdeling. Ik begin met de samenwerking, en wel met de eenvoudigste gevallen, waarbij sprake is van één relatie, dus van één mededeling en één hierdoor opgewekte reactie. Het eerste probleem, wat om oplossing vraagt, is: wat drijft de actor bij het geven van zijn signaal? | |
1. De actor.Wanneer wij nagaan, wat bij ons mensen ten grondslag ligt aan het geven van een signaal, dus aan het ‘iets zeggen’, dan vinden wij drieërlei oorzaken. Wij kunnen soms ‘dom’ reageren op een uitwendige prikkel, bijv. wanneer wij ‘au!’ schreeuwen wanneer wij een verwonding oplopen. Wij kunnen ook op inwendige prikkels met een uitroep reageren. Zo zullen wij eveneens ‘au!’ roepen als reactie op een plotselinge hevige buikpijn. Of we komen tot neuriën of zingen als gevolg van een zeker gevoel van welbehagen of geluk. Tenslotte is er de voor ons belangrijkste oorzaak, die voortkomt uit de behoefte, een ander iets bepaalds mede te delen. Daarbij echter speelt voor ons het effect der woorden een rol; we verwachten, dat onze woorden voor een medemens iets betekenen en dat hij er op zal reageren, hetzij met daden, hetzij met gevoelens. In dit geval wordt dus onze mededeling veroorzaakt door het inzicht dat wij hebben in het effect van onze | |
[pagina 30]
| |
mededeling. Met deze onderscheiding van drieërlei oorzaken wil ik niet zeggen dat onze mededelingen steeds op één van de drie berusten; stellig werken in vele gevallen de drie oorzaken samen. Zo speelt, zelfs bij onze meest ‘verstandige’ opmerkingen, stellig dikwijls een inwendige, niet redelijke, drang mee.
Wanneer wij nu willen nagaan, op welke wijze dieren tot het geven van hun signalen komen, stuiten wij op een grote moeilijkheid. Men kan, door objectief onderzoek, met zekerheid uitmaken of een bepaalde handeling van een dier door een bepaalde prikkel - zij het een in- zij het een uitwendige - veroorzaakt wordt. Maar men kan niet met dezelfde mate van zekerheid uitmaken of een dier een handeling uitvoert doordat het inzicht in het effect van die handeling heeft. Dat ligt daaraan, dat ‘inzicht’ in de grond der zaak niet iets objectiefs is, maar iets subjectiefs, een belevenis, een gevoel, dat wij slechts bij ons zelf, en, tot op zekere hoogte, bij onze medemensen kennen. Bij ons zelf omdat we het immers voelen; hij onze medemensen omdat zij ons iets over hun gevoelens kunnen vertellen. Bij dieren kunnen wij echter oneindig veel minder over die gevoelens te weten komen, omdat dieren ons nog veel minder dan mensen over hun gevoelens kunnen vertellen. Men kan nu echter wel, in het algemeen, met enige waarschijnlijkheid tot een conclusie over het al of niet bestaan van inzicht bij dieren geraken, door na te gaan of zij tot bepaalde prestaties in staat zijn, die bij mensen voortvloeien uit inzicht, d.w.z. samengaan met de belevenis, inzicht te hebben. Het getuigt bij ons mensen bijv. van inzicht, wanneer wij ook dan een weg weten te vinden, om ons doel te bereiken, wanneer de omstandigheden abnormaal zijn. Ook bij dieren kan dit, naar men algemeen aanneemt, een indicator van inzicht zijn, maar met dat overspringen van mens op dier, met die veronderstelling dat bij het dier achter dat op een doel gerichte gedrag | |
[pagina 31]
| |
onder nieuwe omstandigheden ook inzicht schuilt, halen wij natuurlijk een element van onzekerheid binnen, dat wij bij meer objectieve beschouwingen steeds vermijden.Ga naar eind25 Een verder bezwaar is nu echter, dat wij, ook al zouden wij op ‘inzicht’ en ‘wil tot mededelen’ kunnen wijzen, de hierboven gestelde vraag nog niet beantwoord hebben. Deze luidde immers: waardoor geeft de actor het signaal? En als antwoord op deze vraag zijn conclusies omtrent ‘inzicht’ of ‘wil’ nog weinig zeggend. Ondanks deze voorlopigheid en onzekerheid zit er nu echter toch iets bevredigends in, om na te gaan in hoeverre er werkelijk tussen het gedrag van de menselijke en dat van de dierlijke actor overeenkomst bestaat. Wat de wil tot het doen van een mededeling betreft, wanneer we die opvatten als een van binnen opzwellende drang tot het zich-uiten, hoe primitief die drang dan ook zijn moge, dan moeten wij aannemen dat er zoiets bij vele dieren bestaat, dat dus vele dieren onder invloed van een inwendige drang of prikkel een signaal geven. Er zijn voorbeelden te over. Als in het vroege voorjaar de eerste blauwe reigers op de broedplaats terugkeren, kiezen zij zich elk in de vroege ochtenduren een plaats in de bomen van de broedkolonie, hetzij op een oud nest, hetzij op de plaats waar zij later een nest zullen bouwen. Deze eerstelingen zijn alle mannetjes. Op hun post staande, uiten zij met min of meer regelmatige tussenpozen een luide rauwe kreet. De roep heeft signaalfunctie, want hij beïnvloedt andere dieren, hij lokt nl. ongepaarde wijfjes aanGa naar eind26. Uren lang kan zulk een mannetje staan roepen. Een uitwendige prikkel die hiervoor verantwoordelijk zou kunnen zijn, kan men niet aanwijzen. Hoogstens kan men zeggen: het moet niet te winderig, niet te koud en niet te regenachtig zijn. Zijn die remmen niet aanwezig, dan welt van tijd tot tijd van binnen uit de drang op, zich in die lokroep te uiten. Zulk een van binnen opwellende drang is ook voor andere vormen van signaal-gedrag aansprakelijk. Evenals de reiger hebben andere vogels hun sexuele lokroepen, bij | |
[pagina 32]
| |
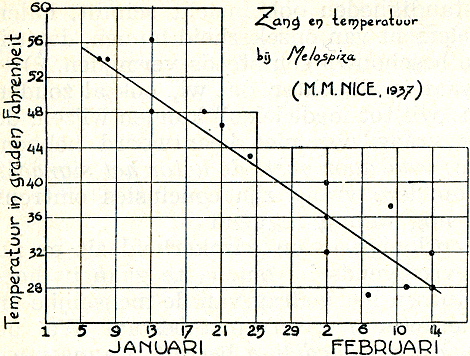 Fig. 17. De veranderlijke afhankelijkheid van de zang van de Amerikaanse zanggors van de temperatuur (Nice). Elke stip is een waarneming van eerste zang na een koude periode.
vele zangvogels bekend onder de naam zang. Dat aan het zingen van de Amerikaanse zanggors bijv. een inwendige drang ten grondslag ligt, blijkt uit het volgendeGa naar eind27. In het begin van het seizoen zingen de vogels alleen bij betrekkelijk warm weer, en houden zich op koude dagen stil. Naarmate nu het seizoen voortschrijdt, hebben koude perioden echter minder invloed. Terwijl bijv. begin Januari een temperatuur van 5 graden C. de zang geheel onderdrukt, heeft dezelfde temperatuur half Februari geen zichtbare invloed (fig. 17). Wij kunnen dit alleen begrijpen als wij aannemen dat er een inwendige, een van binnen uit opwellende drang werkzaam is, die sterker wordt naarmate het seizoen vordert. Hoe later in het seizoen, hoe lager de temperatuur immers zijn moet, om de zang te onderdrukken, m.a.w. hoe sterker de rem, om de steeds sterker geworden inwendige drang in toom te houden.
Beschouwen wij nu de tweede kwestie, die van het inzicht in het te bereiken effect. Bij ons mensen toetst men het inzicht o.a. door bepaalde prestaties in voor het individu nieuwe omstandigheden te eisen, dus door te zorgen dat geen starre, erfelijk of door ervaring vastgelegde, gedragsvorm hem tot het gewenste effect kan leiden. Aan hoe sterker afwijkingen van het gewone de | |
[pagina 33]
| |
proefpersoon zich weet aan te passen, hoe groter mate van inzicht wij hem in het algemeen toekennen. Het is nu al vele malen gebleken, dat reeds betrekkelijk geringe veranderingen in de situatie de meeste dieren tot totaal ‘onjuist’ handelen brengt, d.w.z. tot handelingen, waarmee het doel, het gewone effect, niet bereikt wordt. Een eenvoudig voorbeeld: jonge lijsters sperren, wanneer hun ouders op het nest komen. Dit sperren is, zoals gezegd, een ‘mededeling’ voor de oude vogels, heeft dus signaalfunctie. Wanneer de jongen ongeveer een week oud zijn, wordt hun sperren opgewekt door een visuele indruk van de oude. Bij een analyse van die visuele prikkelsituatie bleek, dat de jongen slechts sperren, wanneer de oude vogel zich voor hen boven ooghoogte bevindtGa naar eind28. Zodra de oude op iets lager niveau was, keken de jongen er wel naar, maar zij sperden niet. Deze kleine wijziging in de omstandigheden (normaliter staan immers de ouden op de rand van het nest en dus ook boven de jongen) was hun reeds te veel. Van inzicht, in de zin als hierboven bedoeld, getuigt dit niet. Het is hier de plaats, om het een en ander over de aard van dergelijke experimenten in het algemeen te zeggen. Men zou bij deze proef met jonge lijsters kunnen tegenwerpen - en men hoort die tegenwerping in zulke gevallen inderdaad dikwijls - dat de dieren hierbij in wel heel onnatuurlijke omstandigheden gebracht zijn. In zeker opzicht is dit juist. Experimenteren is wijzigen van de omstandigheden en deze worden in een experiment dus vanzelf ‘onnatuurlijk’, d.w.z. anders dan de natuur ze biedt. Ware het niet zo, dan zouden we niet hoeven te experimenteren, want dan zou de waarneming onder natuurlijke omstandigheden ons voldoende zijn. Toch is het natuurlijk waar, dat men met het onnatuurlijk maken van de omstandigheden niet te ver mag gaan. Maar waar ligt dit ‘te ver’? De kneep zit, meen ik, hierbij in het volgende. Wijzigt men bij een experiment de omgeving te sterk, dan wekt men bij de proefdieren uitwijk- of vluchtreacties op, | |
[pagina 34]
| |
of men bereikt, dat het dier dodelijk angstig in een hoek gaat liggen. In zulke omstandigheden kan men het dier er niet toe brengen, te gaan voedselzoeken, of te gaan nestbouwen, of te gaan baltsen. Concludeert men dan, uit het achterwege blijven van een reactie op in de proef geboden prikkels, dat deze prikkels van geen invloed op het dier zijn, dan is die conclusie onjuist, omdat onder deze omstandigheden in het dier de vluchtdrang al het andere onderdrukt. Men kan zeggen, dat de omstandigheden te onnatuurlijk waren, maar juister is, te zeggen dat men de juiste verhouding tussen conclusie en experiment uit het oog had verloren. Op zichzelf waren de omstandigheden helemaal niet onnatuurlijk, want ook in de vrije natuur beleeft het dier dergelijke angstige ogenblikken, en als men iets aangaande de vluchtdrang wilde onderzoeken, zou men het dier juist in deze omstandigheden moeten brengen. ‘Onnatuurlijk’ wordt een experiment slechts, wanneer men het dier door te ver gaande wijziging brengt in een andere stemming dan waarin het tijdens de proef behoorde te zijn, dus wanneeer men het dier in vluchtstemming brengt, terwijl men iets over zijn jachtinstincten te weten wil komen, of wanneer men een dier in jachtstemming brengt, terwijl men iets over zijn nestbouwdrang weten wil. Op zichzelf is een proef dus nooit onnatuurlijk, slechts door de aard van de conclusie kan hij onnatuurlijk zijn. Meent men, dat het voorbeeld van de sperrende jonge lijsters nog niet overtuigend is, omdat dit dieren van slechts een week oud betreft, dan wil ik nog een volgend voorbeeld, ook genomen uit de toch betrekkelijk zo hoog staande groep der vogels, geven. Het koekoeksei, dat, zoals bekend, in het nest van een zangvogelpaar door zijn ‘pleegouders’ uitgebroed wordt, komt in den regel op dezelfde dag uit als de eieren van de pleegouders zelfGa naar eind29. De jonge koekoek leeft korte tijd rustig met zijn nestgenoten samen, maar al gauw ontwikkelt hij de neiging, hen er uit te werken. Alles wat hij bij zich in het nest voelt, eieren of jongen, neemt hij op de rug en | |
[pagina 35]
| |
werpt het, achterwaarts kruipend, er
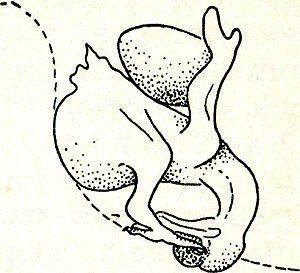 Fig. 18. Hoe een jonge koekoek de eieren uit het nest werpt (Heinroth).
uit (fig. 18). Dit gebeurt dikwijls terwijl een der pleegouders op het nest zit, om eigen kroost en indringer samen te koesteren. Wordt er nu een jong uitgewerkt, dan komt dit min of meer op de nestrand te liggen. Vanaf dat ogenblik ‘herkent’ de oude het ongelukkige dier niet meer als jong, d.w.z. hij negeert het, of werkt het zelfs nog verder weg, zoals hij een op de nestrand gevallen blad zou wegnemen. Geen zijner gewoonlijk op het jong gerichte gedragingen: koesteren, ev. prikkelen tot sperren, voeren, worden verder aan dit jong verkwist, laat staan dat het jong in het nest zou worden teruggelegd. In vermenselijkende taal: hij ziet niet meer in, dat de nieuwe situatie iets met zijn ouderlijke neigingen te maken heeft. Ook hier dus een al heel gebrekkig inzicht. Tot een dergelijke conclusie komt men bij de meeste dieren. Zelfs bij vele zoogdieren, de hoogste uitgezonderd, treft men een zeer lage vorm van inzicht aan. Bij hogere typen kan men van iets dieper gaand inzicht spreken, maar ook deze dieren staan, zoals ik wel niet nader hoef te bewijzen, bij de mens toch heel ver ten achter.
Wij komen dus tot de conclusie, dat er wel zoiets als een wil tot mededelen bestaat, althans in de vorm van een drang om zich te uiten, dat echter van inzicht, althans van een mate van inzicht, voldoende om daaruit de doelmatigheid van de mededeling te kunnen verklaren, in den regel geen sprake is. Ook al zouden wij onze verklaring in deze richting zoeken, dan nog zouden wij al spoedig tot het inzicht komen, dat het bij de dieren anders toegaat dan bij de mensen. Dit betekent de eerste stap in het onderzoek. Wat noopt de actor dan wèl er toe zich te uiten, om zijn signaal te geven? In de eerste plaats dus een drang van binnen uit. Inwendige veroorzakende factoren bewerken, | |
[pagina 36]
| |
zoals we gezien hebben, dat het reigermannetje in het voorjaar gaat roepen, dat de zanggors gaat zingen. We kunnen er aan toevoegen, dat naast inwendige prikkeling ook uitwendige prikkeling een rol speelt: als de reiger in de verte een wijfje ziet, begint hij op slag drukker en luider te roepen. De zang van vele zangvogels staat op een vergelijkbare wijze onder invloed van de waarneming, dus van uitwendige prikkeling; hier doet dikwijls de aanblik van een mannetje, d.i. een territoriumrivaal, de zang losbarsten resp. intensiever wordenGa naar eind30. Ook in andere gevallen weet men, dat in- en uitwendige factoren samenwerken bij het veroorzaken van signaal-bewegingen; dat mededelingen dus tegelijk spontane acties kunnen zijn én door de buitenwereld opgewekte reacties. Soms is het spontane karakter overwegend, bij andere gedragingen het reactie-karakter; dat is van dier tot dier, van signaal tot signaal, van keer tot keer verschillend. Beschouwen we eerst de inwendige drang iets nader. In enkele gevallen weet men iets van de binnen in het dier werkende factoren af en wel ook juist in het geval van de voorjaarszang van vogels. Deze staat namelijk onder invloed van de toestand der geslachtsklieren. Bij kleinere vogels zijn de geslachtsklieren in najaar en winter veel kleiner dan in het voorjaar. Pas tegen de broedtijd beginnen bij het mannetje de zaadcellen zich te vormen, onder grote zwelling van de hele geslachtsklier. Hierbij ontwikkelt zich in dit orgaan een zgn. hormoon, een stof die afgegeven wordt aan het bloed en die door het bloed naar een plaats vervoerd wordt waar een er voor gevoelig orgaan er de invloed van ondergaatGa naar eind31. In het onderhavige geval is het zeer waarschijnlijk, dat het door de geslachtsklier afgescheiden hormoon in causaal verband met de zang, d.w.z. de voor de zang nodige spierbewegingen staat. Hoe het hormoon op het zenuwstelsel inwerkt, dat immers voor het gedrag verantwoordelijk is, weet men niet. Veel opvallender dan het werken van een inwendige drang is dat van uitwendige factoren, en wel van zintuige- | |
[pagina 37]
| |
lijke prikkeling, want bij de hogere dieren
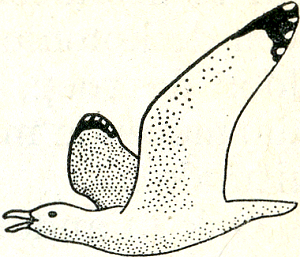 Fig. 19. Alarmerende Zilvermeeuw.
gaan vrijwel alle inwerkingen van buiten via de zintuigen. Het valtimmers, zoals gezegd, dikwijls al bij oppervlakkige waarneming op, dat een dier zich op de een of andere wijzer uit, juist nadat een verandering in de buitenwereld zich voltrokken heeft. Heel duidelijk is dit bv. bij de zg. alarmroepen van vele dieren. Wanneer in een zilvermeeuwenkolonie een hond verschijnt, vliegen de oude meeuwen op (fig. 19) onder het uiten van een luid, rhythmisch herhaal ‘kè’, aldus: ‘kèkèkèkèkè - kèkèkèkèkè!’ Deze roep werkt erg aanstekelijk; alle koloniegenoten raken verontrust en gaan uitkijken naar gevaar, resp, zelf alarmeren. Ook de jongen reageren op de algemene onrust en vooral op de alarmroep, zij verstoppen zich overal in hun vaste schuilplaatsen tussen kruipwilg, helm of duindoorn (fig. 20). De alarmroep der ouden, zonder twijfel dus een signaal (want hij beïnvloedt de gedragingen van andere individuen), wordt door de oude vogels geuit als reactie op het verschijnen van een roofvijand. Die samenhang, tussen het verschijnen van de roofvijand en het uiten van de alarmroep als reactie er op, wordt duidelijk wanneer men het niet bij deze ene waarneming laat, maar het kolonieleven enige dagen lang gadeslaat. Uren lang kan de alarmroep achterwege blijven en alle keren dat hij plotseling overal weerklinkt  Fig. 20. Zich plat drukkend zilvermeeuwkuiken.
vallen samen met het verschijnen van een hond, roofvogel, mens of andere roofvijand. Een dergelijke waarneming, waarbij een proces telkens slechts optreedt onmiddellijk nadat een andere gebeurtenis zich afspeelt, heeft de waarde van een goed experiment, voor zoover het er om gaat, oorzakelijk verband tussen de twee gebeurtenissen te zien. Voor een verdere analyse van de veroorzakende factoren zijn natuurlijk diepergaande experimenten nodig. Zo geeft de waarneming ons weliswaar het recht | |
[pagina 38]
| |
om te besluiten dat het gedrag van de verontruste meeuwen de koloniegenoten alarmeert, maar of dit nu werkelijk door de roep, of ook door de onrustige bewegingen der alarmerende meeuwen komt, kunnen eerst verdere proeven uitmaken. Ook in ander opzicht moet men met het formuleren van een conclusie voorzichtig zijn. Bij de boven al genoemde Natalbaars kan het verschijnen van een roofdier tengevolge hebben, dat de jongen naar de moeder toezwemmen en in haar mond opgenomen worden. Of dit gedrag rechtstreeks het gevolg er van is, dat de jongen de roofvijand waarnemen, of dat ze alleen op de een of andere alarmbeweging van de moeder reageren, valt hieruit echter niet op te maken. Wanneer men deze beperkingen in het oog houdt, kunnen zulke ‘natuurlijke experimenten’, die den onderzoeker in den regel in groot aantal in de schoot geworpen worden, vooral in de eerste stadia van het onderzoek uitstekende diensten bewijzen. Men moet echter op zijn qui vive zijn om de kans waar te nemen zoodra hij komt, want in de schoot valt zo iets alleen wanneer men zich op de juiste plaats weet op te stellen en op het juiste ogenblik actief is. Beschouwen wij nu nog enkele andere voorbeelden, niet zozeer om tot verdere conclusies te komen, als om nog even te demonstreren in hoevele gevallen een signaalbeweging inderdaad zelf re-actie is. Wanneer een stekelbaarsmannetje zijn territorium gekozen heeft, gaat het, zoals gezegd, een ‘nest’ bouwen, een soort tunneltje, van waterplanten geconstrueerd. Zodra dit nest gereed is, verandert de stemming van het mannetje. Zijn kleur wordt anders (het rood feller, de rug lichter) en tevens toont hij plotseling vergrote belangstelling voor wijfjes. Zoodra er een in zijn buurt komt, zwemt hij met een serie van bliksemsnelle, zijdelingse sprongen op haar af en danst als het ware om haar heen (fig. 21). Door deze ‘zigzagdans’ maakt elke stekelbaarsman aan vrouwtjes van zijn soort het hof. Tegenover kuitrijpe wijfjes werkt zijn gedrag ook inderdaad als signaal, | |
[pagina 39]
| |
 Fig. 21. De ‘zigzagdans’ van een roodkaakje voor een drachtig wijfje. (Ter Pelkwijk).
zij voelen zich hierdoor onweerstaanbaar tot het mannetje aangetrokken en zijn bereid hem naar het nest te volgen. Het blijkt nu, door dergelijke waarnemingen als bij het vorige vermeld, dat de zigzagdans zelf een reactie is op het zien van een kuitrijp wijfje. Omgekeerd heeft het wijfje een signaalbeweging die ‘balts’ genoemd wordt, waarbij ze met opgeheven voorlichaam op het mannetje komt toezwemmen. Het mannetje reageert op deze balts door om te keren en het wijfje voor te gaan naar het nest. De vrouwelijke baltsbeweging, die deze reactie in het mannetje opwekt, heeft dus signaalfunctie. Zij is zelf weer reactie op de zigzagdans van het mannetje. Ook deze signaalbeweging heeft dus re-actiekarakter, kan door uitwendige prikkeling opgewekt worden. Dat neemt niet weg, zoals ik hierboven | |
[pagina 40]
| |
al aantoonde, dat ook inwendige factoren van invloed zijn: een wijfje baltst op het zien van een mannetje alléén als ze kuitrijp is; een mannetje bejegent een vrouwtje alleen dàn met de zigzagdans als hij zich in de - slechts weinige dagen durende - phase bevindt die op het voltooien van het nest volgtGa naar eind32. De inwendige en uitwendige veroorzakende factoren spelen meestal een ietwat verschillende rol. De inwendige factoren maken het dier meestal bereid om op de uitwendige factoren te reageren. De inwendige werken voortdurend in; de uitwendige meestal maar even. In gewoon spraakgebruik zou men de uitwendige veroorzakende factoren ‘aanleiding’ noemen. Zulke betrekkelijk eenvoudige signalen kent men ook bij de mens. Het huilen van de baby bijv. heeft immers een heel grote invloed op de moeder; deze voelt de neiging in zich opkomen, het kind te sussen en na te gaan wat zij kan doen om het huilen te doen ophouden. Welk een grote overwinning is het niet voor vele moeders, om een verwend kind ondanks gehuil tòch met rust te laten! Het huilen van de baby, dat dus signaalfunctie heeft, wordt zelf dikwijls opgewekt door uitwendige factoren, zoals kou, of warmte. Gaan wij nu verder na, hoe de actor er toe komt om juist die-en-die bepaalde mededeling te doen, dan komen we een groot verschil met de meeste menselijke mededelingen op het spoor. De mededelingsvormen der dieren behoeven nl. niet zoals onze taal individueel geleerd te worden, maar ieder individu beschikt al van huis uit over de ‘taal’ en omgangsvormen van zijn soort. Zijn vermogen om op het juiste ogenblik het juiste signaal te lanceren zijn aangeboren. Ook bij de mens kennen wij zulke aangeboren mededelingsvormen, men denke weer aan het huilen van de baby, maar zij zijn hier, althans kwantitatief, van ondergeschikte betekenis. Men ontdekt dat signaalbewegingen aangeboren zijn, wanneer men dieren geïsoleerd grootbrengt. Zo heeft men de eieren van stekelbaarzen kort nadat ze gelegd waren uit het nest gehaald en in een afzonderlijk bakje tot ont- | |
[pagina 41]
| |
wikkeling laten komen. Toen de jongen uitgekomen waren, bleven zij van elk contact met soortgenoten verstoken; elk werd in een apart aquarium opgefokt, waar hij nooit een stekelbaars te zien kreegGa naar eind33. Wel groeiden ze ten dele tussen andere vissen op. Een mannetje, dat zodoende een jaar tussen bittervoorns geleefd had, bleef in die hele tijd op en top een stekelbaars en nam geen enkele omgangsvorm der bittervoorns aan. Toen hij geslachtsrijp werd, ging hij een nest bouwen, en toen men daarop een stekelbaarswijfje in zijn aquarium bracht, ontving hij haar met alle bewegingen, te beginnen met de zigzagdans, die een tussen de stekelbaarzen opgegroeide roodkaak tegenover een wijfje ten beste geeft. In zijn gedrag was deze stekelbaars ook voor andere mensen, die het gedrag van de soort goed kenden, niet van andere mannetjes te onderscheiden. Hetzelfde werd bij een vrouwtje vastgesteld; ook zij bleek over alle omgangsvormen van haar soort te beschikken, bleek dus een ‘welopgevoede’ stekelbaars te zijn; contradictio in terminis, want zoals uit dit alles juist blijkt, was elke opvoeding hieraan volkomen vreemd. Zoals het bij de stekelbaars bleek te zijn, zo is het bij vele andere dieren. De gevallen waarin het signaal van andere soortgenoten geleerd wordt, zijn betrekkelijk zeldzaam. De meeste gegevens die wij hierover bezitten, danken wij aan de Duitse onderzoeker Heinroth, die een groot aantal dieren, vooral vogels, van jongsaf aan opgefokt heeft, en daarbij hun ontwikkeling zeer nauwkeurig heeft gadegeslagenGa naar eind34. Volgens Heinroth is de zang van sommige zangvogels, die wij hierboven al als een signaalbeweging leerden kennen, niet geheel aangeboren, maar leren de vogels als jongen aan de zang van de in hun omgeving levende vogels hun eigen zang althans perfectionneren. Meestal zullen die vogels de zang van hun vader overnemen, daar deze immers steeds het dichtst bij is. Blijven de jongen van zulke soorten van elk voorbeeld gespeend, zoals bij Heinroth's dieren wel eens het geval was, dan komt hun zang dikwijls niet verder dan tot een ‘onverstaanbaar’ (niet als van deze | |
[pagina 42]
| |
soort te herkennen) gebrabbel; groeien zij onder andere soorten op, dan nemen zij geheel of gedeeltelijk de zang van vreemde soorten over. Hoe zeer ‘aangeboren’ en ‘aangeleerd’ hier door elkaar heenwerken, blijkt wanneer men jongen laat opgroeien tussen oude vogels van vreemde soorten èn van hun eigen soort. Zij leren dan tòch de zang van hun eigen soort, waaruit toch een zekere aangeboren voorkeur blijkt. | |
2. De rol der zintuigen.Voordat ik nu de bewegingsvorm der signalen verder behandel, moeten wij enkele andere kwesties onderzoeken. Wij zijn gewend, bij het woord ‘taal’ te denken aan de menselijke spreektaal, die wij in ons opnemen door middel van het gehoor. Taal behoeft echter in het geheel niet op acustisch gebied te liggen, want wij spreken ook wel van gebarentaal, en deze is werkzaam op visueel gebied. En in beginsel kan men voor het doen van een mededeling van elk zintuiggebied gebruikmaken, al leent dan ook het ene zintuig zich er veel beter toe dan het andere. Gaan wij dus nu eerst na, welke zintuigen de dieren bij hun ‘taal’ gebruiken. Om uit te maken, op welk zintuiggebied een signaal inwerkt, moet men in de meeste gevallen zijn toevlucht nemen tot het experiment. Zonder experiment kan men weliswaar met enige mate van waarschijnlijkheid tot conclusies komen, vooral als men onder wat wisselvallige omstandigheden waarneemt en men dus de beschikking over ‘natuurlijke experimenten’ heeft, maar men kan zich toch ook lelijk vergissen. Zonder experimenteel ingrijpen zou men bijv. niet gemakkelijk kunnen aantonen dat het sjirpen van sprinkhanen wèl en het zoemen van honingbijen niet acustisch als signaal werkzaam is. Men nam voor het zoemen der bijen dan ook signaalfunctie aan, totdat men langs proefondervindelijke weg ontdekte dat bijen totaal doof zijnGa naar eind35. De proeven die moeten uitmaken welk zintuig van de reactor geprikkeld wordt en zijn gedrag beïnvloedt, zijn | |
[pagina 43]
| |
meestal technisch heel eenvoudig, maar bij de opzet moet enige zorgvuldigheid in acht genomen worden. Men moet nl. om na te gaan, welke prikkeling een dier tot een bepaalde reactie noopt, om te beginnen deze reactie in het dier naar believen kunnen opwekken. Om na te gaan, waarop het stekelbaarswijfje reageert als ze tegen een roodkaak baltst, moet men dat wijfje ‘op commando’ kunnen laten baltsen. Pas dan kan men de uitwendige situatie gaan veranderen en nagaan, bij welke verandering de reactie nog wèl en bij welke zij niet meer optreedt. Deze veranderingen moeten in het ons hier bezighoudende geval bestaan uit wijzigingen, die slechts één prikkelgebied betreffen. Men kan bijv. de chemische eigenschappen van de actor wijzigen zonder hem visueel te veranderen, of hem visueel wijzigen zonder zijn geluid te beïnvloeden. Daar dit zelden mogelijk is door de actor zelf te veranderen (over zoiets als een ‘Tarnkappe’ beschikken wij helaas niet, en de meeste operaties veranderen méér aan het dier dan men wil) moet men de actor eerst vervangen door een kunstmatige nabootsing, een model, een marionet. In deze marionet kunnen wij dan wijzigingen aanbrengen, en nagaan of de reactie er op zich wijzigt. Wil men bijv. nagaan of de rode kleur van een roodkaak een signaal is dat het wijfje tot baltsen prikkelt, dan gaat men na, of de reactie van het wijfje soms achterwege blijft, wanneer men haar een model aanbiedt dat geen rood draagt. Is dit inderdaad het geval, en treedt de reactie wèl op wanneer hetzelfde model wèl rood draagt, dan kan men concluderen, dat de rode kleur invloed heeft, mits men natuurlijk zeker weet dat het roodmaken geen chemische veranderingen meebracht die invloed op het wijfje zouden kunnen hebben. Het zal hier al duidelijk zijn, dat het in zulk een experiment (zoals in alle analytische experimenten) om vergelijken gaat. Men vergelijkt de werking van model A (met rood) met die van model B (zonder rood). De verschillen tussen A en B moeten in de proeven die ons hier interesseren, zoals gezegd, op één zintuiggebied liggen; alle overige eigenschappen (grootte, vorm, geur, | |
[pagina 44]
| |
plaats, bewegingswijze, duur van de proef enz. enz.) moeten volkomen gelijk gelaten worden. Wil men nagaan of het wijfje bijv. op chemische eigenschappen van het mannetje reageert, dan vergelijkt men de werking van twee volkomen gelijke marionetten met elkaar, waarvan de ene alleen in bepaalde chemische eigenschappen van de ander verschilt. De techniek van zulke proefjes ontwikkelt zich tijdens het onderzoek ‘vanzelf’. Een eenvoudige manier bijv. om chemische (reuk- en smaak-) en tasteigenschappen aan een actor te ontnemen, is hem achter glas te zetten. Soms kan dit met de actor in levende lijve gebeuren, zodat men dan nog niet zijn toevlucht tot marionetten behoeft te nemen. Men kan ook, inplaats van de uitzending door de actor te beïnvloeden, de zintuigen van de reactor één voor één uitschakelen. Het mooist is dit natuurlijk, wanneer het zonder schade voor het dier kan gebeuren. Bij insecten bijv., wier hele lichaam met een chitinelaag bedekt is (een dood afscheidingsproduct van de huid), kan men de ogen zonder bezwaar met een laagje zwarte lak bedekken, waardoor ze tijdelijk uitgeschakeld zijn. Zodra de proef voorbij is, kan men de lak weer verwijderen. Het dier wordt hierdoor niet veel meer beschadigd dan onze nagels, wanneer we er lak opbrengen en er weer afhalen. Andere experimenten eisen een meer diepgaand ingrijpen, dat aan den vakman overgelaten moet worden. Men heeft het in deze dingen tamelijk ver gebracht en het is in vele gevallen mogelijk, een zintuig dat men uitschakelen wil, operatief buiten werking te stellen, zonder de levensvatbaarheid van de proefdieren te vernietigen. Zo heeft men blinde vissen, salamanders zonder reukvermogen, dove sprinkhanen enz. lang in het leven kunnen houden in een, op het gemis van een zintuig na, volmaakt gezonde toestandGa naar eind36. Ik wil nu een overzicht geven van een aantal uiteenlopende gevallen die wat deze kwestie betreft min of meer volledig onderzocht zijn. Ten dele grijp ik daarbij terug op feiten, die ik al in het beschrijvende hoofdstuk genoemd | |
[pagina 45]
| |
heb, ten dele zullen er ook weer nieuwe feiten op het tapijt komen. Toch wil ik er vooral nog eens op wijzen, dat deze revue een heel zwakke afspiegeling is van de rijke verscheidenheid van verschijnselen die nog op nader onderzoek wachten. Wat de geluidentaal betreft, deze kennen wij slechts bij enkele diergroepen, en wel bij de geleedpotigen, waar vooral sprinkhanen en krekels mooie gehoororganen bezitten, en bij de gewervelden, waar men een goed ontwikkelde geluidentaal vooral bij de vogels aantreft. 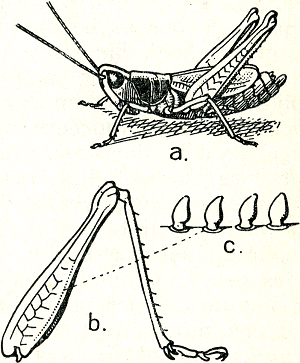 Fig. 22. Het geluidsorgaan van een sprinkhaan. (Weber).
De geluidentaal van krekels en sprinkhanen (fig. 22) is reeds een veertig jaren geleden door de Oostenrijker Regen onderzochtGa naar eind37. Regen merkte, dat de luid sjirpende mannetjes van vele soorten door paarlustige wijfjes opgezocht werden. Hij bood nu in een aantal proeven de geluidsprikkel apart aan, zorgde dus dat het wijfje alleen door het geluid en niet door andere van het mannetje uitgaande prikkels bereikt kon worden, en dat zij het mannetje nòch kon zien, nòch kon ruiken, nòch op een andere wijze kon waarnemen. De fraaiste proef die hij deed bestond daarin, dat hij een mannetje in een kamer voor een telefoon opstelde. Hij bestudèerde dan de reacties van wijfjes, die in een andere kamer gehouden werden, tegenover een in die kamer opgestelde luidspreker, die het sjirpen van de mannetjes weergaf. De reactie was duidelijk genoeg: de wijfjes sprongen in de luidspreker! Voor de functie van aanlokken van grote afstand zijn, ook bij dieren die goed zien, geluiden in gebruik. Voor alle dieren die tussen begroeiïng leven is het immers al op | |
[pagina 46]
| |
vrij korte afstand onmogelijk, elkaar op het gezicht te vinden. Vele vogels vertrouwen dan ook bij het vinden van hun maat meer op het gehoor dan op het gezicht. Zoals boven al gezegd, heeft de zang van zangvogels, juist als de besproken sexuele lokroep van de blauwe reiger, de functie om een wijfje van verre te lokken. Die functie is in vele gevallen zó precies vastgelegd, dat het mannetje het drukst zingt zolang het nog ongepaard is. Zodra hij dan een wijfje krijgt, houdt zijn zang op en gaat hij haar op andere wijze het hof maken. Bij de sneeuwgors heeft men zelfs waargenomen, dat een reeds gepaard en dus zwijgend mannetje opeens begon te zingen, wanneer hij, door onachtzaamheid, zijn wijfje even uit het oog had verlorenGa naar eind30. Deze zang van de zangvogels is alleen mogelijk doordat bij hen een apart stemorgaan tot ontwikkeling gekomen is, een zeer gecompliceerd apparaat, dat op de plaats van samenkomst der twee bronchiën ligt. Deze syrinx, zoals het orgaan heet, ligt dus op een andere plaats dan ons stemorgaan, de pharynx, die immers vrij ver naar voren in de luchtpijp ligt. Er zijn ook vogels, die over geluiden beschikken, die precies dezelfde functie hebben als de zang der zangvogels, maar die wij niet gewend zijn als zang te betitelen, omdat ze ons minder aan menselijk gezang doen denken: het ratelen van de nachtzwaluw, het sjilpen van de huismus en zoals gezegd de rauwe lokroep van de reiger. Het is bij de biologische beoordeling van deze dingen echter tamelijk onverschillig of wij, mensen, één soort uit honderdduizenden, het geluid mooi vinden of niet. Wij moeten al deze sexuele lokroepen, die alle de functie hebben een maat van verre aan te lokken, onder één naam samen vatten, waarvoor het nu eenmaal in gebruik genomen woord ‘zang’ het meest aangewezen is. Men spreekt dan ook zonder bezwaar van de zang van een reiger, van een mus of van een nachtzwaluw. Niet alle vogels zingen met een echt stemorgaan. Er zijn er die op een heel andere, mechanische manier zingen, zonder echte ‘stem’ dus. De bonte specht en vele familie- | |
[pagina 47]
| |
leden van hem lokken hun wijfjes door een
 Fig. 23. Grote bonte specht.
geluid dat ze maken door snel met de snavel op een goed klinkende boomtak te roffelen (fig 23). De zang van een watersnip bestaat uit het zgn. blaten, een geluid, dat door de buitenste staartpennen wordt voortgebracht wanneer de snip zich met extreem gespreide staart steil omlaag stort (fig. 24). Aan dit, sterk op het blaten van een geit lijkende geluid heeft deze vogel de naam ‘hemelgeit’ te danken. Bij andere groepen van gewervelden is de taak van geluiden die op het eerste gezicht wel zang lijken niet zo goed onderzocht. Toch komen ze ook hier veel voor. In de groepen der kikkers en padden bijv. kan men de wonderlijkste soorten van zang te horen krijgenGa naar eind38. In ons land kennen de meeste mensen eigenlijk alleen de bruine en de groene kikker en de gewone pad. De boomkikker echter heeft een nog veel opvallender zang, een luid rollend gekwaak en juist zoals de groene kikker beschikt hij over een resonneerorgaan. Terwijl de groene kikker echter twee resonneerblazen heeft, die bij het kwaken ter weerszijden verschijnen, heeft de boomkikker één grote kinblaas. Een dergelijke reusachtige  Fig. 14. De ‘hemelgeit’: blatende watersnip.
resonneerblaas heeft ook ons gestreepte duinpadje. En komt men in Zuidelijker streken, dan ontmoet men onder deze groep nog veel meer wonderlijke muzikanten. Nooit zal ik de duizendstemmige kikkerkoren vergeten, die de nachten in Florida van hun klanken vervulden: hoge en lage kwaaktonen, waaronder droge, houterige en volle metaalachtig klinkende, zoet gefluit en schril getier, elk geluid typisch voor een aparte soort. | |
[pagina 48]
| |
Zoals ik al gezegd heb, is de functie van de meeste dier zanggeluiden een lokken van de geslachtspartner. Dikwijls echter heeft de zang tegelijk een afschrikkende werking op mannelijke soortgenoten, die er daardoor van weerhouden worden om zich op het territorium van de zanger te wagen. Heel duidelijk is dat bij vogels als de winterkoning en de roodborst. Deze vogels zingen ook 's winters, als er van een wijfje lokken geen sprake is. Wèl verjagen ze dan soortgenoten van hun gebiedGa naar eind39. Geluiden hebben echter lang niet altijd de functie van zang. Vele geluiden zijn zachter en worden op korte afstand gebruikt, om andere reacties te ontketenen: de haan roept zijn kippen bij een lekker hapje; jonge vogels maken bedelgeluiden tegen de ouden die deze tot voedselgeven prikkelen; de leden van een zwervende mezentroep zorgen dat ze elkaar niet kwijtraken door voortdurend op elkaars lokroepen te reageren enz. enz. Natuurlijke experimenten over de rol van het geluid ziet men dikwijls bij in holen broedende vogels. Als men bijv. bij een nesthol met jonge spechten staat en de oude vogels zijn daardoor een beetje angstig, zodat ze niet durven gaan voeren, kan het bedelgeluid van de jongen, die zij immers zelf niet zien, hen opeens tot voeren brengen. Verder gaan bij vele dieren aan de paring geluiden vooraf, die stellig tot taak hebben, de paardrang van de maat te verhogen. Velerlei geluiden dienen verder er toe, om soortgenoten te alarmeren. Kauwtjes bijv. hebben een rauwe, krassende roep die ze aanheffen als een roofdier of mens in de buurt van de kolonie komt, speciaal en heel heftig als die roofvijand een kauwtje te pakken krijgtGa naar eind40. Het effect van dit signaal is, dat alle kauwtjes uit de kolonie op de roepende afkomen en samen met hem de rover gaan bestoken, door hevige duikaanvallen op hem te doen en te proberen hem in staart, rug of achterpoten te pikken. Nòch een wezel of hermelijn, nòch een kat zijn hierop gesteld, en het resultaat van zulk een aanval is dan ook, dat zij ijlings de | |
[pagina 49]
| |
benen nemen, en neiging krijgen, het gebied van de kolonie te mijden. Er zijn ook soorten, die over meer dan één lokroep beschikkenGa naar eind41. Zulke soorten hebben dan ook evenveel reacties, die tegenover verschillende soorten van roofvijanden gebruikt worden. Bekend is dit bijv. bij gewone kippen. Het ene alarmsignaal, een lange, eenlettergrepige, schorre roep is een reactie op een overvliegende roofvogel of kraai; deze roep wekt in de kippen in de omgeving de neiging op, zich plat op de grond te drukken. Tegenover een lopende roofvijand, kat of hond bijv., uiten zij de veellettergrepige, luide alarmschreeuw die zij ook na het leggen van een ei uiten, en de reactie daarop is niet een zich-plat-op-de-grond-druken, maar de neiging om ‘op stok’ te gaan. Men kent dergelijke gevallen bij verschillende andere vogels; de gewone huismus bijv. reageert op een lopend roofdier door het bekende nasale ‘kèw, kèwkèw’ te zeggen, waarop alle mussen naar hem toekomen en zij gezamenlijk boven het roofdier in een struik gaan zitten schetteren, terwijl de reactie op een vliegende roofvogel, een zacht, maar doordringend ‘kruu’, alle mussen tot stilzitten en zwijgen brengt. Wij zijn er nog verre van verwijderd, de vele mogelijkheden van geluidentaal die wij bij dieren verwezenlijkt vinden, goed te kennen. Zelfs bij vogels, waar we wat dat betreft het verst gevorderd zijn, kennen we slechts hier en daar de beteekenis van een geluid. Ik geloof zelfs, dat er geen enkele vogelsoort is, waarvan we het volledige geluidenrepertoire begrijpen; zelfs weten we nog maar van heel weinig vogels precies over welke geluidssignalen zij beschikken. Een grote moeilijkheid daarbij is het registreren van de geluiden op een wijze die de reproductie voor ieder kenbaar maakt. Als een Engelsman het over het geluid heeft dat hij als ‘quaw-eet’ beschrijft en een Duitser over ‘krü-it’, dan is het voor een Fransman waarschijnlijk niet gemakkelijk, om daarin een roep van een nachtzwaluw te herkennenGa naar eind42. En de objectieve registratie van geluiden | |
[pagina 50]
| |
op gramofoonplaten of klankfilm is nog steeds een betrekkelijk kostbare geschiedenis. Toch zijn daarbij al heel mooie resultaten behaald en deze werkwijze zal, ook al doordat zij de mogelijkheid tot experimenteren over de functie dezer geluiden opent (men denke aan de proeven van Regen) nog van grote betekenis blijken te zijn. Zoals ik al opmerkte, is onze kennis bij andere dieren nog veel geringer. Zo weet men bijv. eerst betrekkelijk kort, dat vele vissen uitstekend horen kunnen. Men weet ook, dat sommige vissen geluiden kunnen maken. Maar men heeft er geen flauw idee van, of deze - of andere - geluiden sociale functie hebben. De mogelijkheid bestaat immers ook, dat het gehoor van vele vissen niets met ‘taal’ te maken heeft, maar het dier bijv. op roofvijanden attent maakt. Waartoe dan echter een door de vis zelf geproduceerd geluid zou dienen, moet dan weer op andere wijze verklaard worden. De vissen zijn ook in dit opzicht weer ten zeerste de moeite van het bestuderen waard. Van de geluidentaal naar de volgende soort, de geurtaal, is een grote stap. Want terwijl wij ons met geluiden en zelfs met visuele signalen, zoals gebaren, zelf heel goed verstaanbaar maken, zegt een geursignaal een mens bijzonder weinig. Bij vele dieren echter, bijv. dieren die geen gehoororgaan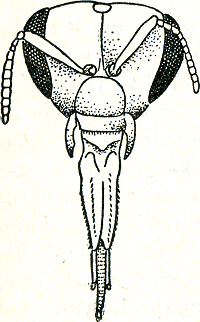 Fig. 25. Kop van een honingbij-werkster (Weber).
bezitten, speelt de geurentaal een grote rol. Eén voorbeeld heb ik al genoemd, nl. dat van de ‘dans’ van de honingbij. De thuiszittende werkeloze bijen, die door dansende pioniers gewekt worden, betrommelen deze van alle kanten met hun sprieten. In deze sprieten bevinden zich, naar men door afzonderlijke proeven vastgesteld heeft, de reukzintuigen (afbb. 25 en 26). Het blijkt nu, dat de bijen, die door de danseres geactiveerd zijn, overal in het hun bekende gebied naar de geur gaan zoeken, die de danseres in haar ‘pels’ met zich meedraagt, en pas gaan honingzuigen als zij een bloem met die geur gevonden hebben. | |
[pagina 51]
| |
Men kan dit met een heel eenvoudige
 Spriet (antenne) van een bij, met reukzintuigjes (von Frisch).
proef bewijzenGa naar eind43. In een tuin waar geregeld bijen komen, stelt men op verschillende plaatsen tafeltjes op. Op elk tafeltje komen een viertal kartonnetjes te liggen, en elk dezer vier kartonnetjes geeft men, door er een druppeltje etherische olie op te sprenkelen, zijn eigen bloemengeur. Elk tafeltje heeft zodoende vier verschillende geurbronnen, bijv. lavendel, thijm, sering en jasmijn. Op elk tafeltje vindt men dezelfde set. Bij één dezer geurbronnen, bijv. lavendel, staat op alle tafeltjes een bakje met suikerwater. Nu wachten de waarnemers, waarvan er bij elk tafeltje één is opgesteld, op de eerste bij die hun opstelling zal bezoeken. Dit kan lang duren, maar eindelijk komt er toch een. Zodra deze zich volgedronken heeft en vertrokken is, waarschuwt de gelukkige waarnemer zijn medewerkers; deze halen overal het suikerwater weg en wachten de dingen af die komen gaan. Het duurt nu niet lang of op alle tafeltjes komen bijen en overal komen ze alleen op het lavendelkartonnetje af en niet op een der andere geuren! De eerste bij was een zoekerbij en heeft de lavendelgeur in de korf meegebracht, waarschijnlijk in de haren hangend; de hierdoor geactiveerde bijen onthouden en herkennen deze geur in het veld. Had men het suikerwaterbakje op het kartonnetje met jasmijn gezet, dan waren overal de jasmijnkartonnetjes bezocht. Een grote rol speelt de geur ook, bij bijen en ook bij mieren, bij het herkennen van de volksgenoten. Leden van | |
[pagina 52]
| |
andere volken, andere kolonies dus van dezelfde soort, worden in het volk niet geduld)Ga naar eind44. Vergist een bij zich bij de oriëntatie en komt hij in een verkeerde kast binnen, dan wordt hij verjaagd of gedood. Hij wordt als indringer herkend aan de ongewone geur die hij bij zich draagt, zijn eigen nestgeur. Bij de stekelbaars, overigens een sterk visueel ingestelde diersoort, vindt men ook een vorm van geurtaal. Wanneer het wijfje door het mannetje naar het nest geleid is, toont hij haar met een bepaalde beweging de nestopening. Hierop kruipt zij naar binnen, wat dikwijls pas na veel moeite gelukt, en zij blijft dan in het nest liggen, de kop aan de ene kant er uit stekend, de staart aan de andere kant. Het mannetje stelt zich nu achter het wijfje op en gaat in snel rhythme met de snuit tegen de staartwortel van het wijfje aanstoten, het ‘siddert’. Tengevolge van dit sidderen legt het wijfje al spoedig haar eieren in het nest af, en kort daarna zwemt zij uit het nest weg. Zodra het Fig. 27. Stekelbaars: een roodkaakje prikkelt het wijfje tot kuitschieten door rhythmisch stoten tegen de staartwortel (Ter Pelkwijk).
| |
[pagina 53]
| |
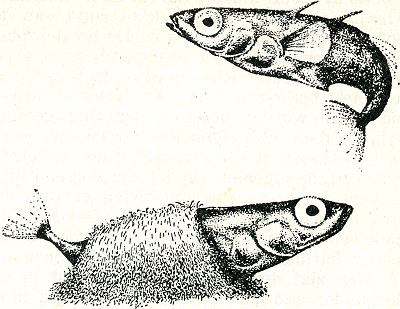 Fig. 28. Het roodkaakje bevrucht de eieren (Ter Pelkwijk).
mannetje nu merkt, dat er eieren afgezet zijn, kruipt het ook in het nest en bevrucht ze (fig. 27). Het is nu gebleken, dat het mannetje daarbij niet op het zien van de eieren reageert, maar op de geur ervan, althans op chemische eigenschappen van de eieren. Wanneer men nl. vers gelegde eieren op een voor hem onzichtbare plek in het nest legt, kruipt hij óók in het nest om ze te bevruchten. Ja, zelfs het vlak bij versgelegde eieren weggehevelde water wekt, als men het in het nest brengt, zijn reactie op. Nauwkeurige visuele nabootsingen van de eieren daarentegen, die men in het nest legt, nopen het mannetje niet tot bevruchten. Zo spreken hier de eieren een taal en wel een chemischeGa naar eind45. Een heel eigenaardige vorm van geurtaal kan men ook bij honden waarnemenGa naar eind46. Ieder kent het dikwijls zo hinderlijke urineren van reuen, vooral op vooruitspringende hoekstenen. En het valt ook steeds op, dat een reu de hoekstenen in zijn gebied elke keer als hij er langs komt besnuffelt. Deze plaatsen zijn voor hem dan ook van bijzondere betekenis: zij zeggen elken vreemden reu, door middel van | |
[pagina 54]
| |
de persoonlijke geur die er van uitgaat, dat hier bezet gebied is. De eigenaar van zijn kant ruikt aan de hoekstenen of er vreemden geweest zijn. De honden planten op deze wijze ‘geurvlaggen’, die op dezelfde wijze als de zang der zangvogels dit op acustische wijze doet, met chemische middelen het bezet-zijn van een gebied aankondigen. Vreemde reuen worden door de geurvlag geïmponeerd; wagen zij het toch, hun eigen urine over zulk een bezette steen te sprenkelen, dan ontwikkelt zich een eigenaardig duel: zodra nl. de eigenaar dit bij zijn volgend bezoek bespeurt, plant hij een nieuwe geurvlag er over heen, en wordt tevens geprikkeld tot herhaald bezoek. Tenslotte loopt zulk een duel uit op retireren van een van beiden; in bijzonder hardnekkige gevallen zelfs op een werkelijk gevecht. Men ziet deze dingen het zuiverst in gevallen, waar de honden aan zichzelf overgelaten zijn; in de stad, waar soms veel honden op een beperkt gebied huizen, en waar de eigenaar in zijn kwaliteit als ‘leiderhond’ dikwijls bepaalt, waar zijn hond verkeert, kan men het systeem niet zo zuiver in werking zien. Waarschijnlijk gaat het ook niet om het afpalen van een individueel gebied, maar van een groepsterritorium. Een soortgelijke wijze van territoriumafpalen heeft men bij hyena's ontdektGa naar eind47. Deze bezitten een klier in de buurt van de anus, die een zalfachtige stof afscheidt. Deze ‘stempelen’ zij op geprononceerde posten, zoals honden dat door besprenkelen doen. Men weet niet zeker, of dit gedrag met territorium samenhangt (de waarnemingen zijn in een dierentuin verricht, en wel in Artis), maar er behoeft m.i. wel nauwelijks aan getwijfeld te worden. Ook marters planten geurvlaggenGa naar eind48. Een heel wat bekoorlijker vorm van geurtaal heeft men ontdekt bij de al even genoemde heivlinderGa naar eind49. In hoofdstuk I beschreef ik, hoe het paarlustige mannetje van deze soort het tegenover hem staande wijfje op ingewikkelde manier het hof maakt. Eerst staat hij te ‘wapperen’; door het rhythmisch openen en weer sluiten van de voorranden der vleugels, waarbij de achterranden gesloten blijven, ont- | |
[pagina t.o. 54]
| |
 II. Eumenis semele (L.), de heivlinder, een onzer inheemse zandoogjes. Links mannetje, rechts wijfje; boven rugzijde, onder buikzijde. Het mannetje draagt op elke voorvleugel een ‘geurorgaan’, op de linker vleugel zwart omlijnd. Naar Tinbergen, e.s., 1943.
| |
[pagina 55]
| |
staat een afwisselend
 Fig. 29. Gewone schubben en een schub uit het ‘geurveld’ van een heivlinderman (B.J.D. Meeuse).
voor- en achterwaarts gerichte luchtstroom, die dus telkens, bij het openen, tussen de voorvleugels van het mannetje weggezogen wordt en, bij het weer sluiten, in de richting van het wijfje naar buiten geperst wordt. Het wijfje strekt gedurende deze ceremonie haar sprieten (die ook bij deze dieren de reukzintuigen dragen) in de richting van het mannetje. Later, bij de buiging, worden de nog steeds voorwaarts gestrekte sprieten van het wijfje tussen de vleugels van het mannetje geklemd. Nu dragen de binnenkanten van de voorvleugels van het mannetje een bizonder orgaan. Over een veld van circa 2 X 12 mm staan hier inplaats van gewone schubben zgn. geurschubben, dunne penseeltjes, welker bouw hen bizonder geschikt maakt tot het snel verdampen van een stof die zich over de haren van de penseeltjes kan uitspreiden (fig. 29). Die stof, die het dier voor het wijfje zo karakteristiek doet geuren, is waarschijnlijk afkomstig van kliertjes die aan de basis van elke geurschub te vinden zijn. Ontneemt men de mannetjes deze geurschubben, dan baat hun al het wapperen en buigen niet veel; de wijfjes laten zich door zulke van hun geur ontdane minnaars niet licht bekoren. Soortgelijke vormen van geurbalts komen stellig bij vele vlinders voor; maar zij zijn maar in weinig gevallen behoorlijk onderzocht. Ook bij andere dieren kan men het bestaan ervan met recht vermoeden, bijv. bij onze gewone watersalamanders. Heeft een mannetje een wijfje ontmoet, dan begint hij met een beweging, die vermoedelijk een | |
[pagina 56]
| |
visuele demonstratie is; hij zet zijn kam op, kromt de rug, maakt zich zo hoog mogelijk en stelt zich in deze houding dwars voor het wijfje op (fig. 30). Daarop maakt hij een formidabele sprong waarbij hij met de gekromde staart
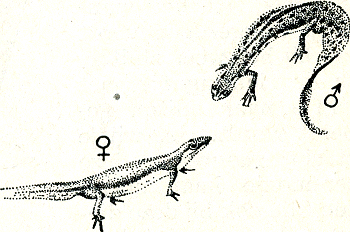 Fig. 30. Kleine watersalamander: het visiuele pronken van het mannetje.
een flinke waterstoot in de richting van het wijfje geeft; dikwijls wordt zij hierdoor van de plaats geduwd. Dadelijk daarop staat het mannetje met het gezicht naar het vrouwtje en begint, door met de naar voren gekromde staart een golvende beweging te maken, een op de kop van het wijfje gerichte waterstroom te verwekken (fig. 31).  Fig. 31. Kleine watersalamander: de ‘geurserenade’ (Ter Pelkwijk).
Deze waterstroom spoelt langs zijn eigen lichaam vóórdat hij het wijfje bereikt; het is waarschijnlijk dat zij hierdoor chemisch geprikkeld wordt, en dat hiervoor speciale geurklieren van het mannetje dienenGa naar eind50. Zo zou ik met het beschrijven van min of meer bekende gevallen door kunnen gaan; ook hier staat het onderzoek nog erg aan het begin. De waterstoot en de er op volgende geleidelijker stroom die de mansalamander op het wijfje afzendt, prikkelen haar wellicht niet uitsluitend chemisch, maar beïnvloeden misschien ook haar tastorganen, al hebben wij daar in het geheel geen bewijs van. Wèl hebben wij bij andere dieren zekerheid over ‘taal’ die van tastprikkels gebruik maakt. Eén voorbeeld noemde ik al hierboven: het sidderen van de roodkaak tegen de staartwortel van het wijfje. Dat hier inderdaad van tastprikkels sprake is, bleek uit proeven, waarbij het lukte het mannetje weg te vangen zodra het | |
[pagina 57]
| |
wijfje in het nest gekropen was, en waarbij vervolgens het wijfje tot kuitschieten gebracht kon worden door haar op de manier van het mannetje met een glasstaaf aan te stoten. Op ingewikkelde tasttaal berust ook de paring van vele slakken. Nauwkeurig onderzocht is dit bij de gewone wijngaardslak, de soort die wij als ‘escargot’ uit de Franse keuken kennenGa naar eind51. De verschillende bewegingen waarmede de parende slakken elkaar stimuleren, bleken alle tastprikkels te leveren, zoals Szymanski kon aantonen, die de juiste reacties in een slak kon opwekken, door met een penseel de passende tastprikkels toe te dienen. Het slot van de paringsinleiding is een wel zeer gewelddadige ‘tast’-prikkel: elk der twee slakken stoot een scherpe kalknaald, de zgn. ‘liefdespijl’ diep in het lichaam van de ander. Spoedig hierop volgt de eigenlijke bevruchting. Een bizonder soort tastprikkel speelt een rol bij de gevechten van vele vissen. Wie bijv. wel eens Indische kempvisjes in zijn aquarium heeft gehad, kent stellig de merkwaardige vechtwijze der mannetjes, waarbij ze zich evenwijdig naast elkaar opstellen en met de staart sterke stoten geven in elkaars richting. Het merkwaardige daarbij is niet zo zeer dat ze met hun staart slaan, als wel dat ze elkaar hierbij niet aanraken. Wèl verwekt zulk een staartslag een flinke stroomstoot in de richting van den tegenstander, en blijkbaar gaat het om die stroomstoot, want deze werkt in hevige mate intimiderendGa naar eind52. Vissen hebben nl. op de lichaamshuid, in de zgn. zijlijn, een zeer gespecialiseerd tastzintuig, waarmee zij bijv. de locale waterstroompjes, zoals kleine prooidieren die opwekken, kunnen waarnemen. Fijne, door een geleiachtig omhulsel omgeven zintuighaartjes dienen er toe, de stroomprikkel op te vangen; zij worden door het bewegende water opzij gedrukt en dat prikkelt de aan hun basis gelegen zintuigcellenGa naar eind53. Een stroomstoot zoals een vechtende vis die geeft moet wel een zeer sterke prikkel leveren. Verdere voorbeelden van tasttaal treft men bij de | |
[pagina 58]
| |
copulatie van vele dieren aan. De copulatie-organen dragen meestal velerlei soorten van tastzintuigen. Ik kom nu tot de bespreking van de categorie visuele signalen. Deze zijn van alle signalen wel de meest opvallende, omdat zij gebruikmaken van bont gekleurde structuren, die dan ook van ouds de morphologen en systematici gepuzzeld hebben. Ze zijn bovendien dáárdoor nog dikwijls bizonder opvallend, dat oogdieren (waarbij we immers juist de visuele signalen kunnen verwachten) door de voortreffelijke oriëntatie waartoe juist hun ogen hen in staat stellen, over een zeer rijk bewegingenrepertoire en dus over vele signaalbewegingen beschikken. Gevallen van visuele signaalbewegingen en -houdingen kent men bij zeer vele dieren, maar slechts in weinige gevallen is hun functie experimenteel onderzocht. Dat ligt aan het feit, dat men met marionetten niet erg goed bewegingen kan nabootsen; en juist de gevoeligheid voor kleine verschillen in bewegingswijze is bij vele visuele dieren bizonder ontwikkeld, niet minder goed, wellicht dikwijls beter dan bij ons. Bekend is bijv. het geval van de ‘rekenende Elberfelder paarden’, die door kloppen met de hoef op de grond juiste antwoorden op allerlei rekensommetjes, zelfs op moeilijke worteltrekkingen, wisten te geven. Na onderzoek bleek het, dat de paarden bij het geven van een antwoord eenvoudig begonnen met rhythmisch met een poot op de grond te slaan, en daarmee doorgingen tot de eigenaar met een haast onmerkbare hoofdbeweging (waarvan hij zichzelf niet eens bewust was) te kennen gaf, dat het nu genoeg was. De hoofdknik was het gevolg van het gespannen meeleven van de oprecht trotse eigenaarGa naar eind54. Zó gering was de beweging waarop het paard zichzelf gedresseerd had, dat men de dresseur niet van de werking er van heeft kunnen overtuigen, nòch hem de beweging heeft kunnen afleren. Degene, die de rekenende paarden heeft kunnen ‘ontmaskeren’, kon hen echter, ten bewijze van de juistheid van zijn conclusie, misleiden door de hoofdbeweging op ongelegen ogenblikken na te bootsen. De paarden hielden dan op met kloppen en maakten fouten. | |
[pagina 59]
| |
Goed onderzochte gevallen waarin de
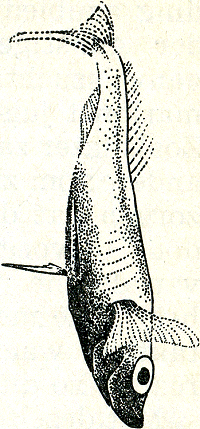 Fig. 32. Dreigende roodkaak.
bewegingsvorm de functie van een signaal heeft, kent men weer bij de stekelbaars. Hierboven beschreef ik de dreighouding der mannetjes: onder opzetten van de stekels en de flank naar de vijand kerend, staan de roodkaakjes bijkans verticaal met de kop omlaag (fig. 32). Men heeft de intimiderende werking van deze houding op de volgende manier bewezen. Dat de werking op visueel terrein ligt, blijkt uit het feit, dat een roodkaak tegen zijn spiegelbeeld dreigt en ook tegen collega's in naburige aquaria. In een bak, waarin twee mannetjes een nest hebben, en ieder dus een eigen gebied heeft, neemt men mannetje A weg. Hierop overschrijdt B spoedig de grens, begeeft zich op A's territorium en plundert diens nest. Zet men nu A terug, dan verjaagt hij B onmiddellijk weer. Men kan echter ook A  Fig. 33. De proef over de dreighouding (Ter Pelkwijk)
in een glazen buisje in zijn territorium terugbrengen, waardoor hij er dus wel is, maar geen aanval kan doen. Hangt men het buisje, dat zo nauw moet zijn, dat A er zich amper in bewegen kan, verticaal, zodat A ‘in dreighouding’ op de kop staat, dan is B merkbaar sterker tot vluchten geneigd dan wanneer men A horizontaal ophangt (fig. 33). Ook kan men dezelfde proef met marionetten nemen, die men in de ene proef in dreighou- | |
[pagina 60]
| |
ding aanbiedt, in de andere proef horizontaal houdt. Onder gunstige omstandigheden, nl. wanneer B niet te aanvalszuchtig òf te bang is, vlucht B voor de dreigende marionet juist wèl, voor de gewoon zwemmende juist niet. Zoals later zal blijken, is de beweging hier één factor naast andere; om zijn werkzaamheid aan te tonen, moet men dus zorgen dat de stemming van de reactor heel labiel is en door toevoeging van juist de ene te onderzoeken factor van de ene reactie in de andere overslaat. De keuze van het juiste moment voor zulk een proef is een kwestie van taktiek, waarmee men moet zorgen, dat het verschil in reactie zo duidelijk mogelijk zichtbaar wordt. Met dezelfde marionet kan men de signaalfunctie van de baltshouding van het wijfje aantonen; het is zelfs wel gelukt om een roodkaak afwisselend tot dreigen en tot leiden te brengen door de marionet om beurten met de kop omlaag en met de kop omhoog aan te bieden. 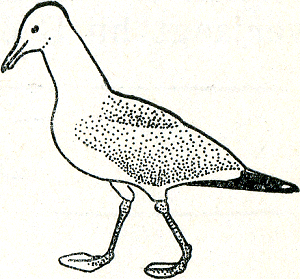 Fig. 34. Dreigende zilvermeeuw.
Over mooie duidelijke signaalbewegingen beschikt ook de zilvermeeuw. Hierboven hebben wij al met de houding kennisgemaakt waarmee een ongepaard vrouwtje een mannetje het hof maakt. Heel dikwijls ziet men ook in een zilvermeeuwenkolonie een agressieve meeuw. Deze stemming uit zich in het schuin omhoog-voorwaarts strekken van de hals, die tegelijk wat stijf en dik gemaakt wordt, en een scherp omlaagwijzen van de kop (fig. 34). Tevens worden de vleugels uit de draagveren geheven. Een dergelijke meeuw ziet er inderdaad imposant en stoer uit, en zijn houding wordt door géén meeuw misverstaan! Wanneer een meeuw vluchten wil, strekt hij eveneens de hals, maar hierbij maakt hij zich niet dik, integendeel, hij legt zijn veren zo vlak mogelijk tegen het lichaam aan (bij alle vogels een voorbereiding voor het opvliegen), en knikt zijn kop niet omlaag, doch eerder ietwat omhoog (fig. 35). Voor de onervaren waarnemer lijken deze twee | |
[pagina 61]
| |
bewegingen veel op elkaar; voor de meeuwen betekenen zij twee uitersten en wanneer men in een meeuwenkolonie door enige seizoenen van nauwgezette waarneming kind in huis is, begrijpt men niet, dat een ander niet onmiddellijk kan zien, of een meeuw ‘boos’ of ‘bang’ is.
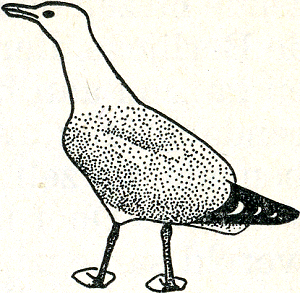 Fig. 35. Zilvermeeuw in vluchtstemming.
Zo is het met vele andere dieren. Bij vele vissen, reptielen, vogels en zoogdieren vooral krijgt men bij geduldig waarnemen al gauw oog voor de vele houdingen en bewegingen en zelfs voor de mimiek der dieren en dan ontdekt men telkens weer nieuwe vormen van bewegingentaal. Vele dierfokkers bereiken in dit begrijpen van de bewegingentaal dan ook het bijna ongelooflijke, en ook bij stropers en jagers treft men wel eens een bijzonder fijne kijk op deze dingen. Vele nog in de ‘natuurstaat’ levende jagersvolken kennen hun jachtdieren ook buitengewoon goed, maar dan ook alleen hun jachtdieren, en eventueel hun huisdieren; alle andere soorten laten hen in den regel volkomen koud. Bizonder boeiend is deze bewegingentaal nu in de talloze gevallen, waarin een signaalbeweging tot resultaat heeft, dat een bepaalde lichamelijke, bont gekleurde structuur, die gewoonlijk vrij verborgen gedragen wordt, plotseling in het volle gezicht wordt gebracht. Wanneer bijv. een eend opvliegt, verschijnt op de gespreide vleugel een bont gekleurd vlak, de zg. spiegel. Deze is bij verschillende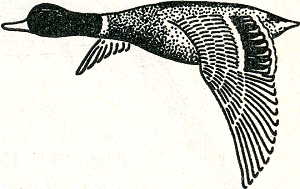 Fig. 36. Wilde eend, woerd.
soorten verschillend van kleur en patroon; het best kent de leek de prachtig blauwe, wit omzoomde spiegel van de wilde eend en zijn in parken veel voorkomende half gedomesticeerde nakomelingen (fig. 36). Deze spiegels dienen als sociaal signaal, dat de soortgenoten in de vlucht aan elkaar bindt, op dezelfde wijze als de lokroepen van in een troepje rondrei- | |
[pagina 62]
| |
zende mezen de troepgenoten bij elkaar houdt. Men heeft in de Berlijnse dierentuin opgemerkt, dat rondlopende eenden en ganzen aan het roepen slaan en op willen vliegen als er eendachtigen overvliegen die eenzelfde vleugelspiegel hebben als zij zelf, ook al zijn zij verder niet zo nauw verwantGa naar eind55. Doordat hier eendachtigen uit alle delen van de wereld samenzijn, kon gemakkelijk vastgesteld worden dat deze reacties alleen van de kleur van de spiegel en niet bijv. van de verwantschap der soorten afhankelijk waren. Bepaalde spiegelkleuren en -patronen, die in hun streek zeer exclusief kunnen zijn, vindt men nl. in verschillende landstreken herhaald en wel bij soorten die helemaal niet nauw verwant met elkaar hoeven te zijn. Bewijzende proeven over de functie van zulke signaalkleuren heeft men o.a. bij een Amerikaans leguaantje Sceloporus undulatus, verrichtGa naar eind56. Dit diertje, zo groot als een gewone hagedis, leeft op de schors van bomen en op rotsachtige droge ondergrond en heeft een prachtige schutkleur. Wanneer een bronstig mannetje op zijn territorium een soortgenoot ontmoet, plaatst hij zich dwars vóór hem, richt zich hoog op de poten op en perst zijn borstkas zijdelings sterk samen, zodat hij hoog en smal wordt (fig. 37). Hierdoor worden grote delen van borst en buik, die eerst ‘onder’-kant waren, plotseling tot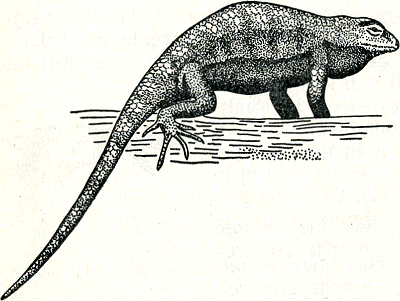 Fig. 37. Pronkende Sceloporus, de ‘fence lizard’. (Noble).
‘zij’-kant en daardoor wordt voor den tegenstander de buikhuid zichtbaar, die onwaarschijnlijk helder blauw gekleurd is. Als het andere dier ook een mannetje is, beantwoordt het dit gebaar op dezelfde wijze, terwijl een wijfje (dat niet blauw van | |
[pagina 63]
| |
onderen is) de beweging niet maakt. Om uit te maken, of dit plotseling demonstreren van de blauwe onderkant signaalfunctie heeft, ging men na, hoe een mannetje reageerde op een ander mannetje, waarvan men de blauwe huid met grijze lak bedekt had en omgekeerd op een wijfje, dat men van onderen en aan de zijkanten blauw geverfd had. In het eerste geval werd het mannetje als een vrouwtje behandeld, d.w.z. de territoriumbezitter probeerde met het proefdier te paren; het blauwgeverfde wijfje echter werd als een mannetje behandeld, d.w.z. aangevallen. De mannetjes beschouwen dus de blauwe onderkant bij elke soortgenoot die ze ontmoeten als een teken van mannelijkheid. Bij de Zuideuropese muurhagedissen, die van onderen eveneens bont gekleurd zijn, rood bijv., staat de zaken ongeveer zoals bij SceloporusGa naar eind57. Iets precieser kent men de functie van de kleur in het vergelijkbare geval van de roodkaak. De rode kleur werkt hier nl. niet alleen tegenover mannelijke soortgenoten als een signaal, maar ook tegenover de wijfjes. Een mannetje dat op zijn eigen territorium is, reageert op een model met rode onderkant zeer vijandig, veel agressiever dan tegenover een blank model. Is hij zelf buiten zijn territorium, bijv. in de situatie van B in de hierboven (blz. 59) beschreven modelproef, dan wekt een rood model veel sterker vlucht in hem op dan een blank model. De vorm van het model doet er bij deze proeven al heel weinig toe; zodra de onderkant van een model maar rood geverfd wordt, bewerkt het intensieve reacties. De rode kleur werkt bij de stekelbaars dus precies zo als de zang bij zangvogels. Een mannetje op zijn territorium valt een zingende man ogenblikkelijk fel aan, terwijl hij zwijgende mannetjes dikwijls duldt. Zodra hij echter zelf op vreemd territorium is, houdt hij zich in het bizonder gedekt wanneer de eigenaar gaat zingen. Tegenover wijfjes werkt het rood van een roodkaak als lokmiddel. Het lukt zelfs om drachtige wijfjes met rode marionetten tot volgen te brengen, ja men kan hen er zelfs toe brengen, zich in een door de marionet aangewezen | |
[pagina 64]
| |
namaaknest naar binnen te werken. Dit lukt echter alleen met marionetten die geheel of gedeeltelijk rood zijn, niet met anders gekleurde. Ook in dit opzicht werkt de rode kleur dus als zang bij vele vogels: afstotend tegenover rivalen, lokkend tegenover wijfjes. Met deze voorbeelden zijn wij dus, bijna ongemerkt, overgegaan van de bewegingen die een opvallend gekleurde structuur demonstreren tot opvallend gekleurde structuren die geen speciale beweging van node hebben om zichtbaar te worden. De rode kleur van een roodkaak werkt op zijn soortgenoten in alle standen waarin het dier zich kan bevinden; hij werkt dus als een permanent tentoongestelde lok- resp. dreigceremonie. Dit uiterste van de reeks: kleur zonder hem ondersteunende beweging, komt echter stellig niet zo algemeen voor als een door een bepaalde beweging naar voren gebrachte structuur. Meestal beschikt een dier over bewegingen, die de structuur zij het dan niet te voorschijn toveren, toch wel iets duidelijker maken. Slechts wanneer de felle kleuren bijna het gehele lichaam bedekken, het hele dier dus eigenlijk één groot signaal is, treden geen speciale demonstratiebewegingen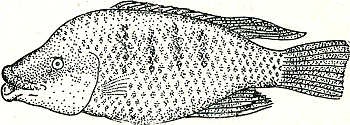 Fig. 38. Natalbaars, mannetje in ‘gewoon’ kleed.
op, hoewel ook bij zulke dieren naar het schijnt dikwijls de neiging bestaat, den tegenstander een groot vlak te laten zien. Bij de roodkaak uit zich dit in het telkens dwars gaan 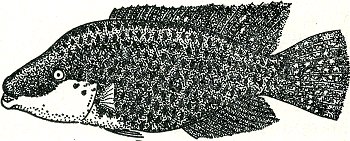 Fig. 39. Natalbaars, mannetje in prachtkleed; het hele lichaam is één groot signaal.
staan tijdens de zigzag-dans en ook tijdens het dreigen. De al meer genoemde Natalbaars wordt in de paartijd als het ware tot één enkel signaal; zijn onop- | |
[pagina 65]
| |
vallende dagelijkse kleur, blank met donkere dwarsstrepen, gaat dan over in een gitzwart kleed; alleen de wangen blijven licht, worden zelfs nog extra wit en de vinnen lopen rood aan (fig. 38 en 39). Visuele signalen maken verder behalve van kleur en beweging ook van vormkenmerken gebruik en sommige signaalbewegingen schijnen speciaal tot taak te hebben, zulke vormkenmerken te demonstreren, zoals andere signaalbewegingen kleurpatronen demonstreren. De baltsbeweging van het rijpe stekelbaarswijfje bijv. lijkt er wel op ‘berekend’ om het dikke achterlijf, teken van kuitrijpheid, aan de roodkaak te demonstreren. En inderdaad blijkt uit modelproeven dat de vorm van het achterlijf voor het mannetje een belangrijk kenmerk is, dat hem zeer sterk tot het uitvoeren van zijn zigzagdans prikkelt. Beschouwen wij nu nog, weer ter demonstratie van de rijkdom der verschijnselen, eens te hooi en te gras een aantal visuele signalen, waarvan de functie door onderzoek bekend is. Wij hebben gezien, dat bij de besproken hagedissen en de stekelbaars, dieren dus waar mannetje en vrouwtje zeer verschillend gekleurd zijn, de dieren het geslacht hunner soortgenoten dus aan de kleur herkennen. Er zijn echter ook soorten, waaronder bijv. vele vogels, waar man en vrouw ongeveer gelijk gekleurd zijn: duiven, meeuwen, vele waadvogels enz. Hier zijn het nu speciale bewegingen die de geslachtsonderscheiding mogelijk maken. Een doffer bijv. bejegent elke duif die hij tegenkomt met de bekende buiging en het spreiden van de staartGa naar eind58. Afhankelijk van de reactie van de tegenspeler hierop (die bij mannetje en wijfje immers verschillend is) wordt deze nu verder als mannetje behandeld of als wijfje bejegend. Heel grote invloed heeft ook bij zulke dieren de houding waarin het wijfje het mannetje tot de copulatie uitnodigt. Het wijfje staat hierbij meestal stil en iets voorovergebogen. Biedt men nu een mannetje van zulk een soort, waar dus mannetje en wijfje gelijkgekleurd zijn, een in die paarhouding opgezette soortgenoot aan, dan volgt dikwijls | |
[pagina 66]
| |
prompt een paringspoging van het
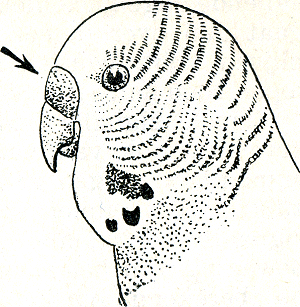 Fig. 40. Kop van een parkiet; de pijl wijst de washuid aan.
mannetje. Het doet er daarbij niet toe of de opgezette vogel een mannetje of een wijfje is; het proefdier merkt blijkbaar geen onderscheid aan de kleine verschillen in grootte, vorm of kleur, die er ook in deze soorten nog wel zijn. Een Amerikaanse onderzoeker die deze stand van zaken bij de Amerikaanse bospatrijs aantoonde, concludeerde hieruit zelfs, dat geen enkele vogelsoort bij de geslachtsonderscheiding op kleurkenmerken zou lettenGa naar eind59. Hierop bewezen twee andere Amerikanen dat dit voor de sexueel dimorphe soorten niet opgaat, en dat bijv. een goudfazantehaan wel degelijk onderscheid tussen zijn opgezette soortgenoten maakt: met een opgezette hen paart hij, een opgezette haan valt hij verwoed aanGa naar eind60. Intussen had reeds lang tevoren een Poolse onderzoekster aangetoond, dat de gewone parkieten elkaars sexe aan betrekkelijk onopvallende kleurkenmerken vaststellen. De washuid van de snavel (fig. 40) is hier bij de mannetjes blauw en bij de wijfjes bruin; verfde zij nu bij een mannetje, dat zij nieuw bij de reeds ingeburgerde (en elkaar dus persoonlijk kennende) parkieten zette, de washuid bruin, dan werd hij als een wijfje door de mannetjes bejegend. Omgekeerd werd een wijfje met blauwe washuid als mannetje bevochtenGa naar eind61. Treffend is ook het gedrag van de gewone inktvis, Sepia officinalisGa naar eind62. Treffend omdat het in allerlei opzichten met dat van sommige vogels en vissen overeenkomt. Ongetwijfeld hangt dat daarmee samen, dat inktvissen, hoe weinig ook met deze beide andere groepen verwant, ook over zeer goede ogen beschikken, waardoor zich ook hier visuele signalen hebben kunnen ontwikkelen. Sepia kan, door samentrekking resp. expansie van duizenden verschillend | |
[pagina 67]
| |
 Fig. 41. Sepia, de zeekat, in rust.
 Fig. 42. Een pronkende zeekat.
gekleurde pigmentcellen in de huid, zeer snel van kleur veranderen. Het ene ogenblik is hij grauw gekleurd als de zandbodem (fig. 41), het andere ogenblik schittert hij met een prachtig patroon van blinkend witte en diep paarszwarte strepen. Vooral het mannetje kan zich prachtig kleuren. Heel duidelijk is die zwartwitte kleuring op twee van de vangarmen, die een beetje breder zijn dan de overige armen (fig. 42). Wanneer een mannetje nu een soortgenoot tegenkomt, (mijn verhaal wordt eentonig), neemt hij de prachtige bonte kleur aan en spreidt de mondarmen zó, dat ze dwars voor de tegenspeler komen te staan, zodat deze ze wel moet zien. Modelproeven hebben ook hier bewezen, dat deze plotselinge demonstratie van een opvallend gekleurde structuur als signaal werkt: de bonte kleur is tegenover de soortgenoten een teken van mannelijkheid. De rode borst van een roodborstje heeft, voor zoover  Fig. 43. Kleurzakjes uit de huid van een inktvis; links uitgebreid, rechts samengetrokken. Om het middelste zijn de spieren getekend, die het kleurzakje uitspreiden.
men weet, een iets andere functieGa naar eind63. Hier hebben man en vrouw beide het signaal, de rode borst, en alleen al daarom zou men niet verwachten, dat het hier precies zoo zou toegaan als bijv. bij de roodkaak. De rode kleur dient dan ook niet als middel ter geslachtsonderscheiding, maar als dreigstructuur. Tegenover elke in het territo- | |
[pagina 68]
| |
rium dringende soortgenoot zet de
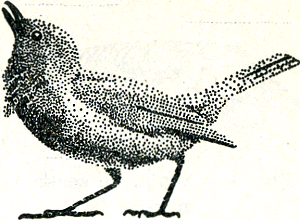 Fig. 44. Een dreigende roodborst (Lack).
roodborst een ‘hoge borst’: de veren worden geheven zodat de rode bef tegenover de omgevende veren sterk naar voren komt springen, en dit grote, iets opgeheven vlak wordt dan de tegenstander toegedraaid, waarbij de eigenaar soms heel gewrongen houdingen moet innemen (fig. 44).  Fig. 45. Goudspecht, vrouwtje.
Bij vele vogels komen aan de kop helder gekleurde vlekken of patronen voor. deze hebben naar het schijnt ook dikwijls signaalfunctie. Het mannetje van de Amerikaanse goudspecht bijv. heeft aan de mondhoek een zwarte vlek, de ‘snor’; het wijfje mist deze vlek (fig. 45 en 46). Door de reacties van een mannetje tegenover een met een kunstsnor beplakt wijfje na te gaan, ontdekte men dat de snor een teken van mannelijkheid is zoals de blauwe washuid bij de parkietenGa naar eind64. Het is waarschijnlijk, dat de goudspecht deze snor bij bepaalde gelegenheden door opzetten der veertjes extra opvallend maakt; hoewel dit nergens beschreven staat, neem ik het aan omdat het bij andere vogels met zulk een snor wel het geval is.  Fig. 46. Goudspecht, man met ‘snor’
Onze Vlaamse gaai bijv., die immers ook een dergelijke zwarte snor heeft, kan hem sterk naar voren brengen. Alleen moet hij hier weer een andere functie hebben dan bij de goudspecht, want bij de gaai dragen mannetje en wijfje die snorvlek. De goudrode streep, die midden over de kop van een goudhaantje loopt, is naar alle waarschijnlijkheid ook een signaalstructuur. Hij wordt tenminste | |
[pagina 69]
| |
 Fig. 47. Dreigende koolmezen (Ter Pelkwijk).
bij territoriumgevechten der mannetjes opgezet en door vooroverbuigingen van de kop aan de tegenstander gedemonstreerdGa naar eind65. De bonte zwartwittekening op de kop van de koolmees is een dergelijk geval. Soms als twee elkaar bevechtende mannetjes tegenover elkaar staan, steken zij hun snavels in de lucht (fig. 47), en draaien in die stand hun koppen zó heen en weer, dat ze elkaar afwisselend de linker- en de rechterwang toekerenGa naar eind66. Of deze ceremonie signaalfunctie heeft, is eveneens niet door proeven uitgemaakt, maar het lijkt alleszins waarschijnlijk. Dit geldt trouwens voor allerlei descriptief reeds bekende gevallen. Bonte kleuren en groteske bewegingen komt men vooral bij vogels en vissen verbazend veel tegen. Wat voor de signaalfunctie van deze verschijnselen pleit, is het feit, dat de bewegingen steeds zo mooi bij de structuren en bij de omstandigheden passen, d.w.z. dat zij er steeds in slagen Fig. 48. Kievitsman, kuiltje drukkend
om de structuren op het juiste moment voor den tegenspeler zichtbaar te maken. De rode kop op het achterhoofd van de gewone kraanvogel wordt gedemonstreerd doordat de vogel zijn tegenstander het achterhoofd toedraaitGa naar eind67. De kaneelbruine staartonderkant van de kievit wordt door het mannetje naar het vrouwtje toegekeerd, als hij haar naar een nest- | |
[pagina 70]
| |
kuil tracht te lokken door hierin
 Fig. 49. Hoe een sneeuwgorsman voor een wijfje pronkt.
te gaan zitten ‘kuiltjedrukken’ (fig. 48). Hij draait het vrouwtje daarbij voortdurend zijn achtereind toe, waar onder de opgeheven staart de helder kaneelbruine verenpartij opblinktGa naar eind68. De al genoemde Amerikaanse goudspecht heeft eveneens een helder gekleurde onderstaartwortel. Hier is hij geel. Om hem aan een soortgenoot te demonstreren, tilt de vogel zijn staart zijdelings op en draait hem tegelijk om zijn lengteas, zodat hij de onderzijde van de staart naar voren richt; de tegenspeler ziet het dus naast zijn opponent opblinken. Merkwaardig is nu weer, dat onze grote bonte specht, die immers rood onder de staartwortel is, deze opvallende verenpartij niet door een zijdelingse beweging demonstreert, maar, o.a. in agressieve ontmoetingen, door de staart onder het lichaam naar voren te brengenGa naar eind69. Een sneeuwgorsman ontvangt een op zijn territorium aankomend wijfje door haar zijn fel wit-en-zwart getekende rugzijde te demonstreren. Hij gaat daartoe met de rug naar haar toe staan, spreidt vleugels en staart iets,; en loopt, over zijn schouder naar het wijfje kijkend, met kleine dribbelpasjes van haar weg (fig. 49). Is hij een meter of wat van haar vandaangelopen, dan komt hij soms nog eens terug en herhaalt de ceremonie. Vele vissen dreigen tegen elkaar door op elkaar toe te zwemmen en tegelijk de kieuwdeksels zo ver mogelijk van het lichaam af te draaien, zodat zij min of meer dwars komen te staan en hun zijkant naar voren gekeerd hebben. Bij sommige soorten zijn nu die kieuwdeksels opvallend van kleur. Leerzaam zijn in dit opzicht twee Cichliden, Cichlasoma meeki en Hemichromus bimaculatusGa naar eind70. Beide soorten dragen op hun kieuwdeksel een diepzwarte, goud omzoomde vlek, die bij het spreiden van de kieuwdeksels | |
[pagina 71]
| |
 Fig. 50. Cichlasoma meeki.
naar voren zichtbaar worden. Bij meeki liggen de vlekken op een andere plaats dan bij bimaculatus, nl. meer naar de buikzijde. In overeenstemming hiermee wordt ook bij meeki het onderste deel van het kieuwdeksel het verste naar buiten gebracht, terwijl bij bimaculatus, waar de vlek veel hoger zit, juist het hoge deel het verst naar voren gedraaid wordt (fig. 50 en 51).  Fig. 51. Hemichromus bimaculatus.
Bij het noemen van deze, tamelijk willekeurig uitgekozen, voorbeelden moet ik het laten. Het zal den lezer duidelijk geworden zijn; dat het werkelijk de moeite moet lonen, om bij elk dier waar we een opvallend gekleurde structuur vinden, naar de signaalfunctie er van te zoeken. Met deze goed gefundeerde werkhypothese in de hand wordt het verblijf in een huidencollectie van een zoölogisch museum ook voor de niet-systematicus een genot. Veel genot beleeft men ook bij het doorbladeren van zoölogische plaatwerken. Telkens ontdekt men nieuwe objecten voor sociologisch onderzoek. Hetzelfde gevoel krijgt men, in nog sterker mate, wanneer men dierentuinen, of soorten- en individuenrijke dierenkolonies bezoekt. Men ziet dan niet alleen de bonte structuren in volle glorie, maar ook de wonderlijke, dikwijls groteske signaalbewegingen in functie, d.w.z. men ziet er maar even een glimp van, maar juist genoeg om te beseffen, dat er een wereld van sociologische problemen op ontdekking en oplossing wacht.
Nu wij dus een aantal signalen hebben leren kennen en in een aantal gevallen weten op welk zintuiggebied ze | |
[pagina 72]
| |
werkzaam zijn, kunnen we verwachten, ook enige algemene kenmerken van signalen te zullen vinden. Bij wijze van werkhypothese heeft men wel eens de volgende gezichtspunten naar voren gebrachtGa naar eind71.
 Fig. 52. Voorjaarsvlucht van een nachtzwaluwman; in de schemering zijn de witte signaalvlekken bizonder duidelijk.
In de eerste plaats zijn signalen steeds opvallend. Daarbij worden vaak zeer interessante middelen toegepast, middelen die de mens soms, zonder hiervoor speciaal bij de natuur in de leer gegaan te zijn, ook toegepast heeft. Meestal wordt de opvallendheid van visuele signaalstructuren bereikt door kleurcontrast tussen signaal en de omgevende lichaamsbekleding. Hierbij worden soms op zichzelf al grote kleurcontrasten door allerlei ‘kunstgrepen’ nog verhoogd, bijv. doordat de grens tussen twee kleurvlakken met een wit lijntje wordt afgezet; men denke aan sommige eendenvleugelspiegels. De kleur van het signaal sluit zich aan bij de waarnemingscapaciteiten van de soort. Zo is het bijv. niet toevallig, dat vogels, die in de schemering actief zijn (uilen, nachtzwaluw) geen bont gekleurde, maar wit-donker getekende signaal-structuren hebben (fig. 52). Bij de lage lichtintensiteit, waarbij hun signalen gebruikt worden, kunnen hun ogen geen kleuren onderscheiden, en daar zijn dus wit-zwart-contrasten het meest geschikt. Een andere manier, om opvallendheid te bereiken, is een bonte structuur afwisselend te laten opblinken en weer verdwijnen, hem te laten flikkeren dus. Dit past de aalscholver toe, als hij bij het ‘vleugelwippen’ de witte vlekken op zijn dijen afwisselend met de vleugels bedekt en weer laat verschijnen (fig. 53). Deze witte lichtreclame is in die zwarte vogel inderdaad heel opvallendGa naar eind72. Een interessante zijde van dit verschijnsel ligt in het | |
[pagina 73]
| |
 Fig. 53. Aalscholver; links in rust, rechts vleugelklappend: de flitsend oplichtende lichtreclame (Kortlandt).
paradoxale feit, dat een opvallende structuur, die weliswaar als signaal zijn nut kan hebben, tegelijk een gevaar in zich bergt, daar het de drager niet alleen voor een mogelijke reactor, maar ook voor een roofvijand beter zichtbaar maakt. Vele signalen worden dan ook verborgen gehouden tot op het ogenblik waarop ze nodig zijn. De middelen waarmee dit bereikt wordt zijn velerlei. In de eerste plaats kan het ‘prachtkleed’ een seizoenkleed zijn. Vele zoogdieren en vogels bijv. ruien op gezette tijden, en krijgen dan aan het begin van de paartijd de signaalstructuren die ze bij de balts en de paringsgevechten gebruiken zullen. Het gewei van herten is een vergelijkbaar geval; allleen schuilt het gevaar hier niet in opvallendheid voor een roofvijand, maar wellicht in de hinderlijkheid. Extreme voorbeelden van die seizoenwisseling zien we bijv. bij eenden, waar de mannetjes in het najaar, hun paarvormingsseizoen, van hun ‘eclipskleed’ dat hun een even mooie schutkleur als de wijfjes geeft, naar hun prachtkleed ruien. Vele dieren, vogels vooral, houden hun signaalstructuren  Fig. 54. Visuele afweerhouding van de zonnereiger (Heilbronn).
ook in het goede seizoen nog min of meer verborgen. Door bepaalde bewegingen kunnen zij dan op het juiste ogenblik te voorschijn gebracht resp. opvallender gemaakt worden. De vleugelspiegels der eenden, de vleugelvlakken van de Zuidamerikaanse zonnereiger (fig. 54), | |
[pagina 74]
| |
de staartpatronen van steltlopertjes (die een soortgelijke functie als de eendenspiegels hebben) worden door spreiden der veren zichtbaar gemaakt. De koplel van de kalkoense haan en andere hoenderachtigen worden met bloed volgeperst en daardoor plotseling zeer veel groter en opvallender. De fregatvogel en de grote trap blazen bepaalde luchtzakken op, waarvan de huid fel gekleurd is (fregatvogel) resp. helder gekleurde veren dragen (trap)Ga naar eind73. Vele vissen bereiken kleurveranderingen met uitzetten of samentrekken van onnoemelijk vele kleurcellen in de huid. Ook inktvissen doen het zoals gezegd op deze wijze. Met al deze maatregelen wordt bereikt dat de kleuring van het dier als geheel een compromis is tussen opvallendheid en onopvallendheid. De signalen moeten echter ook specifiek zijn. Verwarring met signalen van andere diersoorten, en met andere signalen van dezelfde soort, moeten voorkomen worden. Bij visuele signalen wordt dit dikwijls door kleur en vorm bereikt, bij geluidssignalen door melodie en vooral timbre.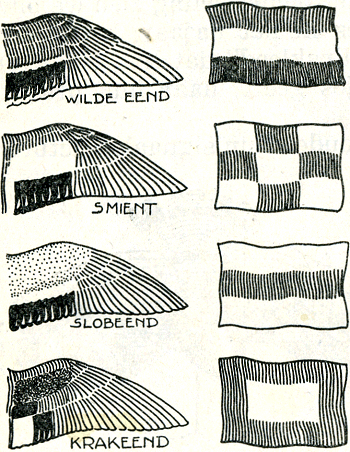 Fig. 55. De vleugelspiegels van eenden wekken dezelfde indruk als vlaggen.
Zelfs bij zangvogelsoorten waar grote individuele variaties in melodie en rhythme van de zang bestaan, kan men de soort dikwijls aan het timbre herkennen. Ongetwijfeld gaat het de vogels zelf net zo. Die specificiteit heeft echter ook weer zijn grenzen. Een ander belang kruist dat van de specificiteit: het belang van de eenvoud. Specificiteit kan men natuurlijk bereiken door middel van zeer ingewikkelde structuren, maar hoe ingewikkelder ze zijn, hoe moeilijker ze op het eerste gezicht van elkaar te onderscheiden zijn. En om herkennen op het eerste ge- | |
[pagina 75]
| |
 Fig. 56. Het bonte scherm van de pauwhaan (bovenstaartdekveren).
zicht gaat het zo dikwijls. Een opvliegende eend gunt zijn rotgenoten niet lang de tijd om te kijken of zijn vleugelspiegel wel die van de soort is. Vooral signalen waarbij het op snel reageren aankomt, zijn dan ook eenvoudig van patroon. Structuren die op een langzaam, summerend stimuleren berekend zijn, zoals de beroemde pauwenveren bijv. hebben wèl ingewikkelde kleurpatronen. Dit samengaan van specificiteit en eenvoud maakt, zoals Lorenz opmerkt, dat een plaat van vleugelspiegels van eenden precies dezelfde indruk maakt als een vlaggenatlas (fig. 55). De mens, verstandelijk werkend volgens dezelfde principes, is tot ongeveer dezelfde oplossing gekomen als de natuur bij de uitrusting van de andere visueel ingestelde sociale dieren.  Fig. 57. Het bonte scherm van de Argusfazantehaan (slagpennen).
Zo zien we ook hier weer een enorme verscheidenheid der ver-  Fig. 58. Het witte scherm van de Bulwerfazantehaan (staartpennen).
| |
[pagina 76]
| |
schijnselen en van de middelen waarmee bepaalde verschijnselen bereikt kunnen worden. Als illustratie van het laatste wijs ik nog op de verschillende wijzen waarop dikwijls verschillende, nauwverwante soorten een zelfde of althans soortgelijk effect bereiken. Van drie hoenderachtigen: Pauw (fig. 56), Argusfazant (fig. 57) en Bulwerfazant (fig. 58) hebben de mannetjes allen het vermogen om bij de balts een opvallend scherm van bontgekleurde veren op te richten. Elk der drie doet het echter met zijn eigen middelen: de Pauw met zijn staartdekveren, de Argusfazant met zijn vleugels (waarvoor het deze, tot vliegen weinig geschikte organen in een hoogst wonderlijke stand moet brengen) en de Bulwerfazant met zijn staartpennenGa naar eind74. | |
B 3. De oorsprong der signaalbewegingen.Wij zijn er nu aan toe, om de vorm der signaalbewegingen nog eens nader te beschouwen. We hebben gezien, dat in vele gevallen de beweging past bij een opvallend gekleurde of gevormde signaalstructuur, d.w.z. deze structuur goed zichtbaar maakt. Daarmee is nog niet alles gezegd: vele signaalbewegingen, onverschillig of zij ‘gebruikmaken’ van opvallende structuren of niet, lijken nl. treffend op bewegingen uit heel andere domeinen van het dagelijkse leven. Dikwijls ziet men dat bijv. bij vechtende dieren. Wanneer twee hanen met elkaar vechten, gebeurt het dikwijls dat ze na een verwoed handgemeen opeens even stil tegenover elkaar staan, in een gespannen gebukte houding, met opgerichte kragen, en dan haastig bewegingen, maken alsof ze iets van de grond oppikken. Hetzelfde doen leeuwerikmannetjes bij gevechten op de grens tussen hun territoria. Het dreigen van stekelbaarzen wordt in zijn meest intensieve vorm eveneens door bewegingen begeleid die precies op eetbewegingen lijken. Het merkwaardige in al die gevallen is, dat het dier niet echt eet; al zijn bewegingen verraden vechtlust en geen eetlust; bij stekelbaarzen heeft men zelfs kunnen vaststellen, dat zij in zulke | |
[pagina 77]
| |
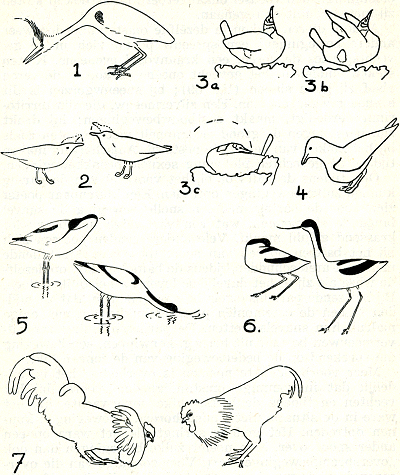 Fig. 59. Verschillende ‘oversprongbewegingen’: 1. hapbeweging van een reigerman (Verwey); 2. bedelbeweging van zilvermeeuwen (N. Tinbergen); 3. vleugelwippen en ‘gorgelen’ van aalscholvers (Kortlandt); 4. ‘hurken’ van een zilvermeeuw (N. Tinbergen); 5. veerpoetsen van een kluut (Makkink); 6. pseudoslapen van een kluut (Makkink); 7. in de grond pikken van hanen (Heilbronn).
| |
[pagina 78]
| |
omstandigheden voedsel onaangeroerd laten, ook al stoten zij met de snuit er tegenaan. Andere dieren maken in dezelf de omstandigheden weer andere bewegingen: een spreeuw poetst zich in de gevechtspauzes de veren, een kraanvogel eveneens. Kluten steken midden in het gevecht opeens de kop in de veren alsof zij gaan slapen (fig. 59); bij sneeuwgorzen is dit eveneens waargenomen. Een zilvermeeuw, die zijn territorium verdedigt, maakt nestbouwbewegingen; hij drukt zijn borst tegen de grond en trappelt met de poten zoals bij het maken van een echte nestkuil. Aalscholvers maken tijdens de gevechten plotseling sexuele pronkbewegingen. Ook tijdens de balts of bij het voorspel van de paring kunnen zulke bewegingen optreden. Een klutenpaar poetst zich voor de paring met een snelle beweging de snavel langs de slagpennen, wat een van verre hoorbaar, dof krassend geluid maakt. Vele andere eenden poetsen zich bij het baltsen op een van soort tot soort verschillende manier. Futen schudden tijdens de balts hun kop op dezelfde wijze als zij in het dagelijkse leven na het duiken doen. Bij baltsende paradijsvogels is waargenomen, dat zij midden tussen de ceremoniën plechtig bukken en met enige nadruk hun snavels wetten. De paringsinleiding van zilvermeeuwen bestaat uit het z.g. kopwippen, een beweging die sprekend op de bedelbeweging van de jongen lijkt. Meer voorbeelden te noemen is overbodig; het is duidelijk dat in sommige omstandigheden, vooral bij het vechten en tijdens de balts, zulke onverwachte, als het ware in de situatie niet thuisbehorende, bewegingen kunnen optreden. Het gedrag springt als het ware op een ander spoor over. Men noemt zulke bewegingen dan ook ‘oversprongbewegingen’Ga naar eind75. Voor sommige van die oversprongbewegingen is vastgesteld dat zij signaalfunctie hebben (nl. voor het dreigen van de stekelbaars en voor het op nestelen lijkende dreigen van de zilvermeeuw); voor vele andere is hetzelfde op zijn minst waarschijnlijk. Er zijn echter ook oversprongbewegingen bekend, die waarschijnlijk geen signaalfunctie hebben. | |
[pagina 79]
| |
Het is opvallend, dat oversprongbewegingen zo dikwijls optreden bij territoriumgevechten, en wel juist op de grens tussen twee territoria. Dit heeft de weg tot een beter begrijpen van de oversprongbewegingen geëffend. Een territorium komt nl. tot stand doordat een dier zijn vechtdrang alleen uit op een bepaald gebied en daarbuiten niet vecht, maar bij een ontmoeting met een tegenstander vlucht. Het resultaat van een ontmoeting tussen twee territoriumbezitters, of het nu vogels, reptielen, vissen, insecten of andere dieren zijn, wordt dan ook steeds bepaald door de plaats waar de dieren zich bevinden; wie op zijn territorium is, heeft de meeste autoriteit (heeft het meeste ‘lef’) en wint het dus. Het is haast uitsluitend een kwestie van bluf, van zelfvertrouwen. Alleen op de grens tussen twee territoria zijn de verhoudingen ongeveer gelijk. Beide opponenten worden hier door het zien van de ander tegelijk geprikkeld tot aanval en tot vlucht. Aanvallen en vluchten tegelijk gaat echter niet. Blijkbaar, zo moet men wel veronderstellen, roept dit conflict tussen aanvalsdrang en vluchtdrang de oversprongbeweging op. Dit wordt vooral waarschijnlijk wanneer wij bedenken, dat ook de mens in zulke omstandigheden moeilijk rustig kan zijn, maar een behoefte aan beweging heeft; hij wordt ‘zenuwachtig’, d.w.z. hij wil bewegen. Men heeft inderdaad experimenteel kunnen aantonen, dat bij de roodkaak de dreigbeweging (die immers over-sprong-eten is) optreedt wanneer het dier tot vluchten geprikkeld wordt terwijl hij in aanvalsstemming is. Deze proef werd als volgt opgezet. Een roodkaak werd in zijn territorium geconfronteerd met een marionet die, door zijn rode onderzijde, zijn aanvalsdrang opwekte. Zodra de roodkaak de marionet aanviel, liet men deze ‘terugvechten’, d.w.z. men stootte de marionet steeds tegen de aanvallende roodkaak aan, zoals een werkelijke stekelbaars ook zou doen. Aanvankelijk werkte zulk aanstoten alleen heftiger aanvallen op, maar men liet de marionet volhouden en dan | |
[pagina 80]
| |
sloeg op een gegeven ogenblik de stemming van de roodkaak plotseling om: hij vluchtte, op zijn eigen territorium, voor de formidabele indringer en kroop in de waterplanten weg. Liet men hem nu met rust, door de marionet kalm te laten hangen, dan kreeg langszamerhand de aanvalsdrang weer de overhand; de roodkaak kwam uit zijn schuilhoek en ging vervolgens vóór het model staan dreigen: de kop ging omlaag en hij stootte met zijn snuit in de zandbodem. Na korte tijd ging dit gedrag weer in daadwerkelijke aanval over. Ging dus de phase, waarin de vluchtdrang overheerste, over in de phase waarin de aanvalsdrang overheerste, dan trad een oversprongbeweging op, juist in de periode dus waarin nòch aanvalsdrang, nòch vluchtdrang de overhand had. Men neemt op goede gronden aan, dat bij zulk een conflict tussen aanvalsdrang en vluchtdrang een drangoverschot in het dier blijft bestaan, dat door middel van bewegingen ‘geloosd’ moet worden. Kan het niet door vechten of vluchten, dan door een oversprongbeweging. Anders lijkt het, althans op het eerste gezicht, te staan met oversprongbewegingen, die tijdens balts of paring optreden. Hier is toch een conflict tussen paringsdrang en iets anders niet waarschijnlijk. Toch moet men ook hier spreken van een drang die zich niet in ‘zijn eigen’ bewegingen kan uiten. Oversprongbewegingen treden hier nl. juist op wanneer de actor onder invloed van zijn sexuele drang staat, maar voor het volledig uitleven van die drang afhankelijk is van een signaal van zijn partner. Wanneer een sneeuwgorsmannetje, dat met zijn vrouwtje wil paren, door haar geweigerd wordt, gaat hij pikbewegingen op de sneeuw maken. De kluutman, die zich voor de paring in de veren staat te pikken, houdt hiermede op wanneer het wijfje opeens de houding aanneemt, die aanduidt dat ze tot de paring bereid is, en die voor hem het signaal is, zonder hetwelk hij de paring niet kan voltrekken. Ook tijdens de balts dus komt dikwijls de situatie voor, waarin een drang, hier de heftige sexuele drang, niet tot uiting kan komen. In dit geval niet door een conflict, | |
[pagina 81]
| |
maar door uitblijven van de noodzakelijke uitwendige prikkeling. Wij zien dus hieruit, dat sommige signaalbewegingen (nl. diegene die oversprongbewegingen zijn) uiting zijn van een hevige drang, die door de omstandigheden niet op de normale wijze uitgeleefd kan worden. Die ‘abnormale’ uiting is dan op zichzelf weer een der aangeboren bewegingsvormen, die het dier ter beschikking staan. Merkwaardig genoeg zijn ook die oversprongbewegingen in elke situatie karakteristiek voor de soort, individuele verschillen bijv. in die zin dat de ene stekelbaars bij een grensconflict eetbewegingen, de andere nestelbewegingen zou maken, zijn er niet. Heel anders is dit bij de mens. Ook de mens maakt oversprongbewegingen, maar die zijn dikwijls individueel verschillend en aangeleerd: de een steekt een sigaret op, de ander speelt met zijn sleutelbos, weer een andere draait aan zijn snor, enz. Het meeste komt echter voor: zich achter het oor krabben. Bij chimpansees is het zich krabben als oversprongbeweging aangeboren. Bij de mens zit in het achter-het-oor-krabben dus vermoedelijk ook een aangeboren element. Nu zijn lang niet alle signaalbewegingen van oorsprong oversprongbewegingen; men kent er ook, die met oversprongbewegingen niets uitstaande hebben. Vogels die bij de langzame nadering van een mens, of door een andere aanleiding, in wegvliegstemming komen, uiten dit door, voordat ze wegvliegen, een paar onvolkomen opvliegbewegingen te maken, meestal bestaande uit hurken, iets heffen van de vleugels, en op- of neergaande kopbewegingen. Men noemt deze bewegingen intentiebewegingen, omdat men er aan kan aflezen wat het dier in de zin heeft. Het schijnt dat intentiebewegingen ook soms signaalfunctie hebben. Men vat bijv. de kopbewegingen die vele eenden maken voordat ze opvliegen als intentiebewegingen op. Bepaalde schuddende bewegingen van de kop, die men bij jongenverzorgende Cichliden kan waarnemen, en die signaalfunctie voor de | |
[pagina 82]
| |
jongen hebben (zij lokken nl. de jongen), zijn eveneens waarschijnlijk intentiebewegingen. Een derde soort van signaalbewegingen zijn de zg. ontwakende bewegingen of symboolhandelingenGa naar voetnoot*. Ook dit zijn onvolkomen handelingen, die echter op een iets andere wijze tot stand komen dan de intentiebewegingen. In het vroege voorjaar ziet men vele vogels dagen, soms weken voordat de volkomen nestbouw begint, onvolkomen nestelhandelingen uitvoeren. Een grasmus-wijfje begint op een goede dag met enkele halmpjes in de snavel te nemen, maar laat ze dan dadelijk weer vallen. De volgende dag draagt ze de halmpjes iets langer mee, de daaropvolgende dag legt ze er een aantal neer in een takvork, weer een paar dagen later bouwt ze een ijl nestbegin, enzovoort, totdat eindelijk, nadat ze op drie, vier plekken zulk een onvolkomen nest gemaakt heeft, een definitief nest tot stand komtGa naar eind76. Futen, ganzen, meeuwen, en allerlei andere vogels sjorren met nestmateriaal voordat er van echt nestbouwen sprake is. Andere vogels beginnen elkaar te voeren lang voordat er eieren, laat staan jongen (waarop het voeren toch eigenlijk ‘gemunt’ is) zijn. Ook broeden ze dikwijls op een leeg nest. Vele maken ook onvolkomen paringsbewegingen in het voorseizoen. Men neemt op goede gronden aan, dat dit alle handelingen zijn die onder invloed van hormonen staan, d.w.z. die pas volledig optreden, wanneer het hormoon in een bepaalde concentratie aanwezig is, of zoals men zegt, wanneer de hormoonspiegel een bepaald niveau heeft bereikt. Ook onder deze ontwakende handelingen kent men er die signaalfunctie hebben. Dit is bijv. stellig het geval bij de paringsinleidingen der ganzen. Beide partners steken hierbij om de 2-5 seconden hun kop in het water en halen hem dan snel weer op. In bizonder intensieve gevallen nemen ze hierbij een bek vol dorre rietstengels e.d. van de bodem op en leggen die met een gracieuze halsbeweging | |
[pagina 83]
| |
naast zich neer. Dezelfde beweging maken zij bij het werkelijke nestbouwen. De dieren gaan hiermee door totdat het wijfje door het innemen van een platte houding, waarbij alleen de hals recht omhoog blijft staan, te kennen geeft dat ze getreed wil worden. Het mannetje stijgt dan op en voltrekt de paringGa naar eind77. De rol van het kopindopen is dus volkomen vergelijkbaar met die van het kopwippen bij de zilvermeeuw of het verenpoetsen van de kluut. Een andere ontwakende beweging is het wederzijdse visjesaanbieden bij sterns. Lang vóór de paring, zelfs voordat de paren gevormd zijn, zweven de visdieven boven het terrein, waar jaarlijks de kolonie huist, rond. Telkens sluiten twee vogels zich even tot een paar aaneen; één heeft een vis in de snavel en vliegt met omlaaggebogen kop achter de ander, die hals en kop ver vooruit strekt (fig. 60). Sierlijk heen en weer zwevend blijven ze zo een tijdje bij elkaar en gaan dan weer ieder huns weegs. Deze Fig. 60. De visjesvlucht der sterns (N. Tinbergen).
onvolkomen beweging ontwikkelt zich later tot echt voeren, welke handeling een sterk bindende invloed op de partners van een paar schijnt te hebben. Het is dikwijls moeilijk, om tussen oversprongbewegingen en ontwakende handelingen te onderscheiden. Het kopindopen der ganzen bijv., hier als ontwakende beweging beschouwd, zou wel eens, bij nadere studie, een oversprongbeweging kunnen blijken. Nadere analyse van de ontwikkeling der beweging en van de omstandigheden waaronder zij optreedt, zal hier uitsluitsel kunnen geven. | |
[pagina 84]
| |
Ook deze feiten zijn weer in tegenspraak met de, boven reeds op andere gronden weerlegde, hypothese, dat een dier signalen zou geven omdat het een mededeling aan een ander wil doen. Er is, dat bleek ook in deze laatste gevallen telkens weer, niet sprake van een iets-willen-meedelen, maar van onwillekeurige bewegingen, die ontstaan doordat opgekropte drang een uitlaat moet vinden (oversprong) of doordat het dier door hormonale resp. nerveuze invloeden tot onvolkomen afreageren genoopt wordt (ontwakende handelingen resp. intentiebewegingen). Het dier kan niet anders dan zich uiten; dat zijn bewegingen, als het ware secundair, óók nog mededeelfunctie, signaalfunctie hebben, gaat hem om zo te zeggen niet aan. Later, in hoofdstuk V, zullen we zien dat er inderdaad meer aanwijzingen zijn, dat we hier van een primaire, de actor ontlastende, functie, en een secundaire, de reactor beïnvloedende functie moeten spreken. Ik moet intussen toegeven, dat de hier besproken conclusies voorlopig slechts voor enkele signaalbewegingen gelden.
Het zal de lezer opgevallen zijn, dat er in deze laatste paragrafen steeds over vogels gesproken is. Dat ligt ongetwijfeld niet daaraan, dat andere dieren minder signaalbewegingen zouden bezitten, althans het ligt niet uitsluitend daaraan. Voornamelijk komt het doordat vogels in dit opzicht zo veel beter bestudeerd zijn dan andere dieren. Bij vissen, waaraan men eerst kort geleden meer aandacht is gaan besteden, blijken zulke signaalbewegingen minstens even talrijk te zijn als bij vogels, en aangezien vissen betere studie-objecten zijn dan vogels (zij zijn gemakkelijker te fokken en voor experimenten te gebruiken) kunnen wij in de naaste toekomst een verdieping van onze kennis vooral van zulke vissenstudies verwachten. Ongetwijfeld echter zullen ook andere diergroepen bij nadere kennismaking vele in dit opzicht interessante signaalbewegingen blijken te bezitten. | |
[pagina 85]
| |
4. Het effect op de reactor.Wij zijn nu gekomen tot het stellen van de vraag: hoe moeten wij ons de uitwerking van de mededeling op de reactor voorstellen? Wat wordt er door de mededeling in de reactor bewerkt, en waardoor bewerkt de mededeling iets in de reactor? Al hierboven werd duidelijk, dat de reactor in vele gevallen onmiddellijk doet, wat zijn naam aangeeft: hij toont een reactie, een antwoordbeweging. Op het horen van de alarmroep der ouden drukt een jonge meeuw zich onmiddellijk tegen de grond. Het lokken van de haan doet de kippen dadelijk naar hem toe rennen. De aanval van een roodkaak jaagt een indringer op de vlucht. Het sperren van jonge zangvogels noopt de oude tot voeren. Het signaal wekt dus in deze gevallen altijd een handeling op, een onmiddellijk, en altijd hetzelfde, antwoord. Als wij hier daarom van ‘antwoordhandeling’ of reactie spreken, wordt daarmee natuurlijk niets gezegd over de mate van inzicht of doelbewustzijn waarmee de reactor in zulke gevallen zou handelen. Men heeft, onder de indruk van die gevallen waarin de reactie onmiddellijk op het signaal volgt, de signalen wel eens aangeduid met de naam ‘Auslöser’ (Engels: ‘releaser’). Een goed Nederlands woord is hiervoor niet in gebruik, en uit wat nog volgen zal, zal blijken, dat het woord signaal wel zo bruikbaar is. Bij nadere beschouwing blijkt namelijk, dat een signaal lang niet altijd een onmiddellijk optreden van een bepaalde antwoordbeweging veroorzaakt. De kwestie blijkt ingewikkelder te zijn dan men op het eerste gezicht zou vermoeden. In de eerste plaats komt het heel dikwijls voor, dat de reactor pas reageert na herhaalde prikkeling, na herhaald geven van het signaal. De zigzagdans van de roodkaak is hier een voorbeeld van. Het is betrekkelijk uitzondering dat een wijfje onmiddellijk op een zigzaggend mannetje toezwemt; meestal moet het mannetje een hele serie sprongen maken voordat het wijfje reageert. Het boven reeds beschreven ‘sidderen’ van de roodkaak is een soortgelijk geval. Dit sidderen, dat een rhythmisch herhaald stoten is, dus een opeenvolging van afzonderlijke stoten, tegen de | |
[pagina 86]
| |
staartwortel van het wijfje, stimuleert haar tot kuitschieten. Meestal moet het mannetje tientallen malen, soms honderden malen tegen de staart van het vrouwtje stoten, eer dit tot kuitschieten komt. Iets ingewikkelder, maar in principe op gelijke wijze, komt de copulatie van zilvermeeuwen tot stand. Aan de eigenlijke paring gaat een hele serie van eigenaardige wippende bewegingen van de kop vooraf, het ‘kopwippen’. Het wijfje prikkelt door deze serie van bewegingen haar man er toe, tenslotte op te stijgen; het mannetje brengt tegelijkertijd door zijn kopwippen het wijfje er toe, bij de paring mee te werken. Op eendere wijze bewerkt het herhaalde veerpoetsen bij een klutenpaar dat hier aan de paring voorafgaat, dat beide partners tenslotte de paring voltrekkenGa naar eind78). Ook in gevallen waarin normaliter éénmaal uitvoeren van de signaalbeweging voldoende is, kan het voorkomen, dat de reactie achterwege blijft en dat pas een herhaling van dezelfde signaalbeweging het antwoord opwekt. Zulke gevallen tonen, dat het signaal de eerste keer weliswaar geen als beweging uiterlijk zichtbare uitwerking had, maar dat het toch iets in de reactor bewerkt had, want dezelfde prikkeling had de laatste maal een ander effect dan de eerste maal. Men kan dat uitdrukken door te zeggen dat de reactor door het schijnbaar tevergeefs gegeven signaal toch al iets ‘in de stemming’ gebracht was. De uitwerking van het signaal blijft enige tijd bestaan, en voordat die nawerking weer genihileerd is, komt het volgende signaal er als het ware bovenop; de uitwerking wordt ‘gesummeerd’. Meestal echter is de situatie nog iets ingewikkelder. Wanneer de na een succesvolle verkenning thuiskomende zoekerbij haar gemoed door het uitvoeren van een honingdans uit, raken de werkeloze bijen in grote opwinding, betrommelen de boodschapster met hun sprieten, vliegen tenslotte uit en gaan op zoek naar de geur die de boodschapster bij zich droeg. Vinden ze een bloem van diezelfde geur, dan strijken ze er op neer, zoeken en verzamelen de | |
[pagina 87]
| |
honing, vliegen er mee naar huis, geven de honing af, en vliegen weer uit. Ze zij nu voor een dansende bij in de korf totaal ongevoelig geworden. Het zou een beetje simplistisch zijn, als we er in dit geval tevreden mee zouden zijn, te zeggen dat de boodschapster niets meer dan een ‘antwoordbeweging’ had opgewekt. Immers wat de uitvliegende bijen gaan doen, is hoogst gecompliceerd: zij zoeken eerst rond, richten zich dan op de hen prikkelende geur, reageren bij het bloembezoek op de honingmerken en op de honing zelf, prenten zich de vorm en de kleur der bloem in het geheugen, vinden door middel van een hoogstaande oriëntatievorm de weg naar de korf terug, enzovoort. Er is dus iets heel anders gebeurd dan het ontketenen van één reactie, en het is juister te zeggen: een heel actiesysteem is in werking getreden. Er is in de reactor iets veranderd, waardoor het hele ingewikkelde samenstel van zoekgedrag, instincthandelingen en oriëntatiemechanismen, al of niet door leerprocessen beïnvloed, dat in de wachtende bij werkeloos gelegen had, in functie is getreden. Nog duidelijker komt dit tot uiting in het volgende voorbeeld. Wanneer in een rustig deel van een zilvermeeuwenkolonie een in de verte geuite alarmroep (‘kèkèkèkè’, het ‘kokkelen’) doordringt, is de reactie van de verschillende meeuwen heel verschillend. Er zijn meeuwen, die op het horen van het alarm dadelijk opvliegen en eveneens gaan alarmeren. Er zijn er ook, die haastig opstaan van het nest, naar een naburig topje lopen, uitkijken en dan alarmerend opvliegen. Weer anderen staan aarzelend op en blijven boven hun eieren staan. Sommigen ook gaan wel even uitkijken, maar keren weer terug naar hun nest en blijven broeden zolang het gekokkel ver weg blijft. Ook zijn de reactie van een en dezelfde meeuw op verschillende ogenblikken niet gelijk. Zo heeft men bijv. waargenomen, dat een meeuw door de alarmroep van een bepaalde buurmeeuw steeds tot opvliegen en meekokkelen geprikkeld werd, doch op het alarmeren van een andere buurmeeuw steeds tamelijk onverschillig bleef. Al zulk | |
[pagina 88]
| |
soort waarnemingen wijzen er op, dat elke meeuw door individuele ervaring langzamerhand weet, uit welke hoek en door welke buurman het kokkelen op ernstig gevaar duidt, en in welke gevallen het zo'n vaart niet loopt. Met andere woorden, ook hier wekt het signaal (de alarmroep) niet één reactie op, maar hij brengt de reactor in een toestand, waarin een bepaald deel van het actie-systeem geactiveerd is. Zulk een toestand noemt men, naar analogie van wat men door zelfwaarneming te weten komt, een stemming. Daarbij wordt vanzelfsprekend niets gezegd over de gevoelens die het dier beheersen; deze kunnen we immers toch niet meevoelen. ‘De meeuw is in alarmstemming’ is niet anders dan een omschrijving van het feit, dat in hem een bepaalde drang geactiveerd is, die hem tot de beschreven gedragsvormen brengt. Inplaats van ‘stemming’ zegt men ook wel eens ‘physiologische toestand’. Deze conclusie lijkt, na de verleidelijk simpele ‘Auslösertheorie’ (volgens welke het signaal een van te voren klaarliggende reactie ‘auslöst’) op het eerste gezicht wat teleurstellend. Het leek zoo mooi: een dubbeltje (de Auslöser) in de automaat (reactor) en er kwam een reep uitrollen (de reactie). Er komt echter geen reep uitrollen, maar een kwispelend hondje, dat naar een reep gaat zoeken en deze, waar hij hem ook vindt, aan degene die het dubbeltje gaf komt brengen. Wanneer wij weten dat het dubbeltje ergens in de automaat electrisch contact maakt, waardoor een rood lichtje aangaat dat het hondje waarschuwt, dan begrijpen wij weliswaar iets van de processen die ons die reep bezorgen, maar er blijft nog heel wat te onderzoeken, wij staan voor het raadsel van het hondje, want wat speelt zich allemaal in dat dier af? Ongeveer zó doet het ons ook aan, wanneer wij weten dat een signaal bijv. langs visuele weg de reactor in een bepaalde stemming brengt, want dan stuiten wij immers weer op de vraag, wat die stemming dan eigenlijk in het dier activeert. De gedragsleer is in dit probleem nog niet heel ver doorgedrongen. Wij zullen ons voorlopig met deze conclusie moeten tevreden stellen, en ons realiseren dat zij | |
[pagina 89]
| |
een begin van onderzoek markeert, eerder dan een eindpunt. Behalve het activeren van een stemming kan ook het onderdrukken van een stemming het gevolg van het geven van een signaal zijn. Een mooi voorbeeld daarvan is door Lorenz bij de kwak ontdekt, een kleine reiger, die op de kop een opvallende signaalstructuur draagt in de vorm Fig. 61. Kwak, de kopveren aangelegd.
 Fig. 62. Kwak, buiging en vertoon der kopveren.
van een donkerblauwe kap met drie draadvormige witte veren, die in rust als één dun wit bandje mediaan over die kap liggen. Komen twee kwakken dicht bij elkaar, dan hebben ze de neiging elkaar aan te vallen. In de regel vermijden ze elkaar dan ook. Wanneer een mannetje en een wijfje samen broeden echter, en dus elkaar op het nest moeten aflossen, en ook wanneer zij samen bij de jongen op het nest moeten zijn, moeten zij wel dicht bij elkaar komen. Een bepaald signaal zorgt er dan echter voor, dat de aanvalsstemming onderdrukt wordt. De arriverende kwak maakt namelijk tegenover de ander een buiging, waarbij hij de drie witte veren heft en tegelijk spreidt, zodat ze als een grote witte hanepoot vóór de blauwzwarte kap komen staan wuiven en dus een heel opvallend visueel signaal vormen (fig. 61 en 62). Het vertoon van dit signaal onderdrukt in de tegenspeler diens agressieve stemmingGa naar eind79.
Wij hebben dus nu gezien dat een signaal de stemming van de reactor beïnvloedt. Soms is het gedrag van de reactor onder invloed van die stemmingsverandering heel ingewikkeld en variabel, zoals in het geval van de meeuwenalarmroep; soms is het eenvoudig en bestaat het werkelijk uit simpele reacties, zoals in het geval van het | |
[pagina 90]
| |
sperren van de jonge zangvogels. Bizonder opvallend is daarbij, dat de reactor zo dikwijls niet individueel reageert, maar steeds op dezelfde wijze als zijn soortgenoten in dezelfde omstandigheden doen of zouden doen. Een drachtig stekelbaarswijfje reageert op de zigzagdans met de baltsbeweging. Alle stekelbaarswijfjes ‘doen dat nu eenmaal zo’. Dat wijst er al op, dat we in zulke gevallen te maken hebben met erfelijk vastgelegde handelingsnormen. Slechts betrekkelijk zelden, en dan nog vooral bij hogere dieren, krijgen we met een sterke individuele variatie, en met afwisselend gedrag bij één individu te maken. Dat die bewegingen in zovele gevallen aangeboren zijn, weet men dan ook inderdaad uit talrijke proeven, waarbij men dieren geïsoleerd heeft opgevoed. De hierboven al vermelde stekelbaarzen waren, zoals reeds gezegd, in gedrag van hun wilde soortgenoten niet te onderscheiden. Vele gevallen, vooral bij vogels, zijn beschreven door Heinroth. Merkwaardig is verder ook de speciale gevoeligheid voor de signalen van de soortgenoten. Terwijl de zigzagdans van een roodkaak op bittervoorns zonder de minste uitwerking bleef, bracht hij een wijfje dat nog nooit een soortgenoot gezien had prompt tot reageren. Die speciale gevoeligheid is dus evenals de handelingsvorm erfelijk vastgelegd. Dus niet alleen het vermogen tot het uitvoeren van de antwoordhandelingen, maar ook de speciale gevoeligheid voor de door het signaal bewerkte prikkeling is erfelijk vastgelegd. Dit geldt nog algemener dan de conclusie omtrent het aangeboren zijn van de handelingen, want ook in gevallen als ik hierboven beschreef, en waarvan het gedrag van de reactor individueel sterk kon verschillen, is de gevoeligheid een kwestie van aanleg die voor alle individuen van dezelfde genetische constitutie gelijk is. Al reageren de verschillende zilvermeeuwen zeer wisselvallig op het kokkelen der soortgenoten, toch is de alarmstemming bij allen het gevolg van het horen van de alarmroep. Géén meeuw laat het kokkelen onverschillig, alle komen er door in alarmstemming, en niet in voedselzoekstemming of in sexuele stemming. | |
[pagina 91]
| |
5. De aard der zintuigprikkels.Dit feit zij aanleiding om de gevoeligheid voor de zintuigelijke prikkeling nog eens nader te onderzoeken. Dat wil zeggen, wij gaan, om in de beeldspraak te blijven, het in de ‘automaat’ geworpen dubbeltje eens op de keper beschouwen. De optimale prikkelsituatie is nog maar in betrekkelijk weinig gevallen grondig onderzocht. Toch zijn voor zulke onderzoekingen objecten genoeg voorhanden; iedere aquariumliefhebber bijv. heeft objecten te kust en te keur. Bovendien kunnen zowel de waarnemingen als de modelproeven met heel eenvoudige middelen uitgevoerd worden. Hierboven heb ik al beschreven, hoe de vechtlust van een stekelbaarsman vooral opgewekt wordt door de aanblik van een andere man van zijn soort. Wij zagen bovendien, hoe met modelproeven uitgemaakt kon worden, dat vooral de rode kleur van den tegenstander als een ‘rode lap op een stier’ werkte: rode modellen worden het heftigst aangevallen. Zolang wij niet meer weten dan dit, komen wij in de verleiding te menen, dat de oplossing van het probleem nu gevonden is. Men zou namelijk kunnen concluderen, dat ‘rood’, ‘iets roods’, de beslissende factor was. Dat wil zeggen dat de cellen van het netvlies slechts een bepaalde hoeveelheid rood licht zouden behoeven te ontvangen om de antwoordhandeling te ontketenen resp. de stemming in het dier te activeren. Een bepaalde hoeveelheid prikkeling van een bepaalde soort, dat lijkt inderdaad op de eigenschappen van het dubbeltje: bepaalde doorsnede, bepaald gewicht, die voldoende zijn om de automaat te laten functionneren. In deze verleiding, om de werking van het signaal met die van het dubbeltje te analogiseren, komt men echter alléén wanneer men bij deze, eigenlijk ietwat slordige, formulering van de conclusie blijft staan. Nadere beschouwing toont nl. al dadelijk, dat de totnutoe beschreven proeven helemaal niet het recht geven te besluiten dat het alleen maar om ‘iets roods’ gaat. Wij weten alleen, dat een model met rode onderzijde (fig. 63) de vechtstemming veel sterker activeert dan een model zonder rood | |
[pagina 92]
| |
aan de onderzijde. Vergelijken we
 Fig. 63. Roodkaakjesmarionet.
nu de werkzaamheid van het eerste model met die van een model dat op de rug rood is inplaats van op de onderzijde (fig. 64), dan blijkt dat dit nieuwe model het mannetje vrijwel koud laat! Dat 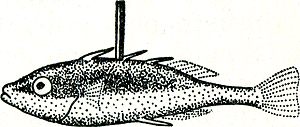 Fig. 64. Roodkaakjesmarionet met het rood op de rug.
bewijst dus dat de optimale prikkelsituatie niet beschreven kan worden als ‘iets roods’, maar als ‘iets dat rood van onderen is’, dat dus het rood in een bepaalde relatie ten opzichte van andere lichaamsdelen moet staan. Het doet er daarbij (binnen zekere grenzen) niet toe, of het model klein of groot is, ver weg of dichtbij. Het doet er dus niet toe, of vele netvliescellen door rood licht getroffen worden of weinige. Bovendien doet het er niet toe, of het aangevallen model sterk verlicht wordt of zwak, ook of het boven den aanvaller of onder hem, links, rechts, vóór of zo ongeveer achter hem zwemt. Met andere woorden, nòch de hoeveelheid rood licht, nòch de plaats of het aantal er door geprikkelde netvlieselementen doen ook maar iets ter zake. De prikkel kan dus niet zomaar gemeten worden, als een hoeveelheid van iets beschreven worden. De werking van die ‘prikkel’ is dus wel een heel andere dan die van het dubbeltje in de automaat. Er zijn in de laatste tijd proeven verricht, die nog verder in deze richting wijzen en die ons dus nog verder van de beeldspraak van het dubbeltje verwijderen. Ik heb beschreven dat de paring van de heivlinder met een reactie van het mannetje op een voorbijvliegend wijfje begint. Naar modelproeven uitgemaakt hebben, is voor het mannetje de kleur en de grootte van het wijfje van betrekkelijk ondergeschikte betekenis, maar het is tamelijk belangrijk welke vorm een model - het wijfje - heeft. Biedt men namelijk twee modellen aan, die een even grote oppervlakte hebben, maar waarvan de verhoudingen | |
[pagina 93]
| |
verschillend zijn, bijv. een rechthoek van 5 × 8 cm en een van 2 × 20 cm, dan krijgt de eerste rechthoek, welks proporties meer met die van een vlinder overeenkomen dan die van de lange rechthoek, verreweg de meeste reacties. Dus ook hier kan men weer niet zeggen dat de prikkel een bepaalde quantiteit van iets moet leveren, maar het gaat om een relatie van lengte en breedte. Deze weinige feiten - onze kennis is hier nog gering - mogen tonen, dat we hier op een moeilijk probleem stoten en er verre van zijn, de werking van de uitwendige prikkeling goed te begrijpen. We zullen ons echter niet verder op het gebied van de zintuigphysiologie resp. waarnemings-ethologie begeven. Eén kwestie moet echter toch nog even genoemd worden. Dat het signaal zo dikwijls niet als een eenvoudige hoeveelheid meetbaar is, maar eerder door bepaalde relaties werkzaam is, sluit niet uit, dat de aangeboren gevoeligheid van de reactor dikwijls zeer speciaal het Fig. 65. Vereenvoudigde roodkaakjesmarionetten met rode onderkant, en een volledige maar bleke marionet. Slechts de twee bovenste marionetten worden door een roodkaak als rivalen beschouwd.
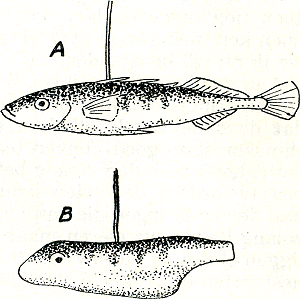 Fig. 66. Stekelbaarsmarionetten; een schematische visvorm met dik achterlijf (B) heeft sterker uitwerking op het mannetje dan een volledige, maar magere stekelbaars (A).
| |
[pagina 94]
| |
signaal zelf geldt, onder uitsluiting van allerlei andere kenmerken. Het rood van het roodkaakje is zó belangrijk dat men het met alle mogelijke vormkenmerken kan combineren, zonder de reactie te beïnvloeden. Modellen van allerlei vorm wekken vechtlust op, als zij maar rood van onderen zijn (fig. 65). Heel duidelijk is ook die eenzijdige, geconcentreerde aandacht voor de signaalstructuur bij het mannetje als hij tegenover een drachtig wijfje zigzagt. Hier gaat het om een signaal dat door zijn vorm werkzaam is. Zó uitsluitend let het mannetje dan op de vorm van het wijfje, die haar kuitrijpheid aangeeft, dat hij meer en intensiever zigzagt op een zeer grof nagebootst visje met een dik achterlijf, dan op een gave, tot in détails echte stekelbaars die een gewoon slank achterlijf heeft (fig. 66). | |
6. Beperking door leerprocessen.Het is nu tijd om te wijzen op de veranderingen die soms tijdens het leven van het individu in de aangeboren gevoeligheid optreden. Door leerprocessen worden nl. de relaties tussen de individuen nog ingewikkelder, en van dergelijke leerverschijnselen kent men een aantal hoogst interessante voorbeelden, die ik nu wil behandelen. Aan den bezoeker van een broedkolonie van meeuwen of sterns dringt zich dikwijls de vraag op, hoe het mogelijk is dat de oude vogels elk hun eigen nest resp. jongen bij thuiskomst zo goed kunnen terugvinden. Wat het nest betreft, heeft men deze vraag betrekkelijk eenvoudig kunnen beantwoorden: de ouden kennen de plaats precies. Men kan de eieren tegen die van andere individuen verwisselen; zolang de nieuwe eieren maar in het goede nest komen te liggen stoort de meeuw zich aan de ruil niet en gaat op het nest zitten broeden. Zó sterk hangen ze zelfs aan de plaats, dat ze er heen terugkeren en er telkens willen gaan zitten broeden, zelfs wanneer men het nest met eieren en al bijv. een meter of twee verplaatstGa naar eind80. Zolang de jongen klein zijn, blijft deze oriëntatie nog bestaan, zij het in minder extreme vorm. Verwisseling met even oude kuikens van | |
[pagina 95]
| |
soortgenoten stoort de ouden in het geheel niet; zij verzorgen elk jong, dat de juiste leeftijd heeft, zonder onderscheid des persoons. Dit verandert sterk als de jongen een paar dagen oud zijn. Verwisselt men hen dàn met even oude vreemde jongen, dan willen de ouden de ondergeschoven kinderen niet accepteren, integendeel, ze pikken hen dood. Het blijkt dat de ouden hun jongen persoonlijk kennen, d.w.z. dat hun aangeboren gevoeligheid voor de signalen die van de jongen uitgaan, door een leerproces vernauwd is, zodat hun ouderlijke handelingen alleen nog maar door de van de eigen jongen uitgaande signalen opgewekt kunnen worden. Heel mooi is dit ook te zien bij de grote sterns, wanneer zij in Augustus in grote troepen op het strand leven, vliegvlugge jongen en ouden dooreen. Zoals bij zovele soorten met heel gespecialiseerde wijze van voedselzoeken (al stootduikende een vis te vangen, zoals sterns doen, is een buitengewone prestatie!) blijven ook bij de sterns de ouden de jongen lang na het uitvliegen nog onderhouden. In Augustus blijven dan dikwijls troepen van honderden jongen op het strand rusten, terwijl de ouden de zee naar vis afzoeken. Komt er een oude met een visje terug, dan vliegt hij naar de troep en gaat luid roepend er boven rondkruisen, al heen en weervliegend, alsof hij iets zoekt. Dat doet hij dan ook stellig. Overal waar hij verschijnt springen de jongen al bedelend in de lucht; sommigen vliegen hem zelfs achterna. Plotseling schijnt hij zijn keuze te bepalen. Met een abrupte zwaai keert hij om, en maakt aanstalten om neer te strijken. Dit kan door het wilde toelopen van jongen nog één of enkele malen mislukken, maar nu laat de vogel zich niet meer afhouden en hij daalt tenslotte en voert een jong. Helaas is het, zolang zulk een waarneming niet bij persoonlijk herkenbare dieren verricht wordt, niet mogelijk om met zekerheid te bewijzen, dat deze vogel zijn eigen jong uitkoos. Er kan echter amper aan getwijfeld worden, omdat deze persoonlijke binding voor verscheidene sterns wel is vastgesteld voor de periode waarin de jongen nog niet kunnen vliegen. | |
[pagina 96]
| |
Nog duidelijker wordt die persoonlijke binding als de jonge sterns iets ouder zijn en zelf ook al wat gaan vissen, zonder daarom de buit, die de ouden hun brengen, te versmaden. Jong en oud zwerft dan, op zoek naar vis, elk op eigen gelegenheid. Heeft de oude iets gevangen, dan vliegt hij luid roepend naar de plek waar hij zijn jong het laatst gezien heeft. Hier vliegt hij aldoor roepend kriskras rond, over een gebied van honderden meters doorsnede. Sommige van de in de buurt vissende jongen schenken geen aandacht aan hem, anderen ook roepen terug of komen zelfs op hem af. Maar de oude blijft roepen totdat één bepaald jong hem antwoordt. Hij schijnt de stem goed te kennen, want onmiddellijk verandert zijn gedrag. Hij daalt langzaam, het jong gelegenheid gevende bij hem te komen, en dan vindt op het water of op de grond de voedering plaats. Hierop vertrekt de oude weer, en ook het jong gaat weer zijns weegs. Bij zulk een gelegenheid kon ik ook eens vaststellen, dat een oude bij drie achtereenvolgende gelegenheden inderdaad driemaal hetzelfde jong voerde. We hoeven er dan ook niet aan te twijfelen of al deze waarnemingen getuigen van persoonlijke bindingen, dat wil zeggen van een door een leerproces specifieker geworden aangeboren gevoeligheid. Hoe dit specifieker worden in zijn werk gaat, zal ik in hoofdstuk V bespreken, waar de ontwikkeling van sociale structuren te berde wordt gebracht. Bij vele vogels herkennen ook man en vrouw elkaar persoonlijk. Heel mooi kan men dat bij zilvermeeuwen waarnemen. Allerlei gedragingen toont een gepaarde meeuw alléén tegen zijn maat. Eens was ik er, in een schuilhutje gezeten, getuige van, hoe een broedend mannetje, wiens vrouw uit voedselzoeken was, bezoek kreeg van een vreemd wijfje, dat hem het hof ging maken. Dat gaat bij zilvermeeuwen nl. van het wijfje uit. De reactie van het mannetje was hoogst interessant. Wel verre van op de avances van het wijfje in te gaan, strekte hij de hals (een teken van alarm) en toen zij dicht genoeg bij hem kwam, hakte hij een paar keer wild naar haar. | |
[pagina 97]
| |
Dikwijls kan men ook in een zilvermeeuwenkolonie zien, hoe een dier, dat van verre komt aanvliegen, zich midden in een grote troep zilvermeeuwen laat vallen, en wel juist naast die meeuw, die bij nader onderzoek zijn maat blijkt te zijn. Het is telkens weer interessant om daarbij te merken, hoe buitengewoon scherp het waarnemingsvermogen van zulke dieren is. Ik heb de bewijzen er van gezien, dat een zilvermeeuw in dergelijke situaties zijn maat in de vlucht op minstens 25 meter kan herkennen, ook als deze niet roept! Dergelijke dingen kent men ook van kauwtjes en grauwe ganzen; bij zoogdieren bijv. ook bij honden, en mogelijk komen persoonlijke bindingen ook bij lagere gewervelden voor. Bij vissen bestaan hiervoor wel aanwijzingen, maar voor zover mij bekend, geen bewijzen. Ieder aquariumbezitter kan hier prachtige ontdekkingen doen. Nog verbazingwekkender is het feit, dat niet alleen mannetje en wijfje, maar ook individuen die verder van elkaar afstaan elkaar zo goed kennen. Dat de kippen in een koppel, de apen in een clan, de Eskimohonden in een sledespan elkaar goed kennen, behoeft na wat ik over de daar heersende rangordeverschijnselen gezegd heb, wel geen betoog. Bij zilvermeeuwen ontdekte ik zulke persoonlijke connecties tussen buren, en wel daaraan, dat een bepaald paar niet alleen zijn eigen territorium, maar ook dat van zijn buren tegen vreemden verdedigde. Nooit echter verdreven deze dieren die buren zelf van dat gebied. Zij maakten dus duidelijk onderscheid tussen hun buren en de andere meeuwen. Soortgelijke feiten beschrijft Kirkman over de kokmeeuwGa naar eind81. Ook aan andere kleinigheden merkt men, dat er tal van persoonlijke bindingen bestaan. Ik heb meeuwen gezien, die tegenover bepaalde buren zeer prikkelbaar waren en bij het minste geringste juist op hèn afstoven. Ook komt het voor, dat een bijzonder licht alarmerende meeuw door zijn omgeving niet au sérieux genomen wordt; ieder weet als het ware: ‘dat is die kwast weer, daar hoeven we ons niets van aan te trekken’. | |
[pagina 98]
| |
Ook de gezusters en broeders uit één gezin leren elkaar dikwijls persoonlijk kennen. Interessante gegevens hierover heeft bijv. Brueckner verzameld tijdens een grondige studie die hij van de sociologie van kippen maakteGa naar eind82.
Hoe herkennen zulke dieren elkaar nu? Bij mijn weten is hierover slechts bij vogels het een en ander met zekerheid bekend. Hier berust het herkennen, naar we trouwens zouden verwachten, op visuele indrukken, of op geluidsindrukken, of - wat ongetwijfeld het meest voorkomt - op een samenspel van beide. Een herkennen aan de stem vond stellig plaats in de volgende gevallen. Een vischdief zit op het nest te broeden. Overal om hem heen zitten zijn koloniegenoten. Er is een constant va et vient van sterns en meeuwen over hun hoofden, onder voortdurend geroep vliegen er dieren naar zee, en komen andere, met vischjes in de snavel, weer terug naar de kolonie. Onze stern blijft kennelijk van al dit lawaai onberoerd, hij zit doodstil met de kop in de wind, en telkens vallen zijn ogen toe. Tenslotte is hij vast in slaap. Opeens, als er weer een roepende stern komt aanvliegen, schrikt hij op, is meteen klaarwakker, en begint zijn bedelroep te uiten, onderwijl rondkijkend. Ogenblikkelijk vestigt hij zijn ogen nu op de naderende stern. Deze blijkt zijn maat te zijn; hij strijkt neer en biedt onze stern een vischje aan. Het lijdt geen twijfel of het roepen van zijn maat had hem gewekt, terwijl al het roepen van de tientallen, misschien honderden, overgetrokken koloniegenoten hem niets gezegd had. Dit ligt ook niet daaraan, dat zijn maat een andere roep, een ander signaal gegeven had dan de andere sterns, want alle met vis thuiskomende sterns uiten dezelfde roepGa naar eind83. Persoonlijke eigenschappen van de stem geven hier blijkbaar de doorslag - het individuele timbre, dat bij elke roep tòch karakteristiek blijft. Precies zo gaat het immers bij ons mensen ook; ook wij herkennen familieleden en vrienden aan hun stem, onafhankelijk van wat zij zeggen. | |
[pagina 99]
| |
Tamme vogels herkennen dikwijls hun menselijke pleegvader eveneens aan de stem. Kleedt men zich wat ongewoon aan, dan zijn de dieren aanvankelijk in de war; roept men hen dan, dan leggen zij op slag hun schuwheid af. Betrouwbare waarnemingen hierover zijn bijv. bekend bij reigersGa naar eind84 en bij de raaf. Indirecte aanwijzingen over het belang van de stem krijgt men ook als men ziet hoe een visuele verandering in de actor het gedrag van de reactor soms in het geheel niet beïnvloedt. Zo hebben Lorenz en ik eens enkele ganzenkuikens, die door hun ouders zeer goed individueel herkend werden, met roet pikzwart gemaakt, een ander met meel wit gemaakt, weer andere met papiersnippers beplakt zodat hun vormen totaal veranderden. Er was bij de ouders zowel als bij de gebroeders enige verwondering te merken, maar alle zo toegetakelde jongen werden toch als eigen jongen aanvaard. Daar zij voortdurend hun infantiele lokroep lieten horen, werden zij vermoedelijk, wel aan hun stem herkend. Bij het reeds genoemde onderzoek van Brueckner bleek, dat kippekuikens hun eigen moeder ook in het donker van andere kloeken wisten te onderscheiden, en wel aan de stem. Op het gezicht herkennen speelt echter stellig ook een grote rol. Ook zwijgende zilvermeeuwen herkennen elkaar met één oogopslag, evenals kauwtjes en ganzen. Bij de sledehonden bestaat daarover ook niet de minste twijfel. Een waarneming van Heinroth bij zwanen maakt het verder waarschijnlijk dat deze dieren elkaar vooral aan de kop herkennenGa naar eind85. Eén zijner zwanen viel nl. eens zijn eigen maat aan, toen deze, met de kop onder water, ‘grondelde’. Toen de aangevallene zijn kop boven water bracht, hield de aanvaller zich opeens in en sloot zich weer vreedzaam bij zijn partner aan. Herkennen aan de kop, aan de gelaatsuitdrukking, wordt ook waarschijnlijk als men bij nauwkeurig toekijken merkt, dat hier inderdaad grote verschillen bestaan. Deze vallen ons in den beginne net zo weinig op als ze ons bij mensen van een ander ras opvallen, | |
[pagina 100]
| |
maar in de omgang krijgt men ongemerkt kijk op de verschillen. Wie wel eens onder een natuurvolk verkeerd heeft, weet hoe eerst alle personen hem identiek voorkomen en hoe hij later niet meer kàn begrijpen dat hij ‘totaal niets op elkaar lijkende’ personen met elkaar heeft kunnen verwarren. Sommige vogelwaarnemers hebben het in het herkennen van hun dieren ook ver gebracht; zij zien verschillen, waar de minder geschoolde ondanks de grootste oplettendheid ze niet kan ontdekken. Bewonderenswaardig zijn in dit opzicht ook de prestaties van vele oppassers in dierentuinen, ook van sommige veeboeren, die hun dieren goed uit elkaar kennen. Toch blijven de prestaties van al deze mensen bij die van sommige vogels en zoogdieren stellig nog achter, tenminste wat het herkennen van dieren betreft. Waarschijnlijk is het zo, dat de mens het best andere mensen herkent, en dat elke diersoort tegenover zijn eigen soortgenoten over die scherpe waarneming beschikt, en bijv. tegenover de mens weer in het herkennen van mensen achterstaat, dus dat iedere soort het gemakkelijkst zijn eigen soortgenoten van elkaar kan onderscheiden. Verschillende waarnemingen wijzen er verder op, dat de hogere sociale dieren zich elkaar persoonlijk lang herinneren. Lorenz beschrijft pakkend, hoe een zijner kauwtjes, na een afwezigheid van zeven maanden in de kolonie terugkerend, door alle koloniegenoten terstond herkend werd, wat o.a. uit de plaats die ze hem in de rangorde weer toekenden en uit het feit dat hij dadelijk geduld werd, duidelijk was.
De kwestie der persoonlijke betrekkingen is, al moet ik hier de behandeling staken, nog in het geheel geen afgesloten hoofdstuk. Dat blijkt reeds uit de fragmentarische behandeling, waarmee ik me tevreden moet stellen; het zal ook nog blijken in hoofdstuk V, waar de ontwikkeling der persoonlijke betrekkingen op het tapijt zal komen. Er ligt hier een hoogst aantrekkelijk studieveld braak, dat niet alleen in sociologisch, maar ook in psychologisch opzicht | |
[pagina 101]
| |
verband tussen onze kennis van de mens en de biologie zal leggen. | |
C. De arbeidsverdeling.Beschouwen wij nu het tweede aspect van een gemeenschap, de differentiatie, nader. De verdeling van arbeid berust zoals gezegd op verschillen tussen de individuen. De vraag is nu, van welke aard die verschillen zijn en hoe, resp. waardoor zij ontstaan. In sommige gevallen zijn die verschillen heel duidelijk, namelijk wanneer zij de bouw der dieren betreffen. De bijengemeenschap bestaat uit koningin, werksters en darren, en de verdeling der functies tussen deze groepen komt in hun bouw duidelijk tot uiting. De koningin is de enige bij in het hele volk, die goed ontwikkelde en functionnerende eierstokken heeft; ze mist echter weer de ‘korfjes’ aan de achterpoten, dat zijn een soort borstels die de werksters gebruiken bij het verzamelen van stuifmeel. De eierstokken der werksters zijn, hoewel in aanleg aanwezig, toch niet tot volle ontwikkeling gekomen. De darren nemen aan het leven der werksters geen deel; zij bezitten evenmin als de koningin verzamelkorfjes en hebben als enige taak, de jonge koninginnen te bevruchten. In verband met het feit dat zij deze in de lucht, tijdens de ‘bruiloftsvlucht’, moeten vinden, hebben zij bizonder goed ontwikkelde ogen. Fig. 67. Een mierenwerkster (links) en een mierensoldaat (rechts).
Bij de andere statenvormende insecten zijn zulke verschillen tussen de ‘kasten’ dikwijls nog opvallenderGa naar eind86. Bij verscheidene mierensoorten vindt men nl. niet alleen koninginnen, werksters en mannetjes, maar onder de werksters komen nog verschillende typen voor. Een zekere bekendheid heeft het type dat men ‘soldaten’ noemt, werksters met enorm grote kaken (fig. 67). Deze ‘soldaten’ zijn inderdaad bij vele soorten de | |
[pagina 102]
| |
vechters van het volk, die bij botsingen met andere volken (bij ‘rooftochten’ bijv.) het vechtwerk doen; in andere gevallen is de betiteling ‘soldaat’ minder juist. Bij vele termietensoorten treft men eveneens soldaten aan (fig. 68). Daar zijn het echter niet alleen wijfjes, maar ook mannetjes, zoals trouwens elke kaste der termieten uit mannetjes en wijfjes bestaat. De kaste der soldaten is bij vele termietensoorten weer in onderkasten verdeeld, waarvan de kaken weer verschillend van grootte en vorm zijn. Over de functie dezer verschillende soldatentypen weet men nog niet veel; ook hier is het niet zeker of wel alle typen met grote kaken werkelijk vechtfunctie hebben. Voor sommige vormen met staafvormige, kurketrekkersgewijs gewonden kaken lijkt twijfel aan de vechtfunctie gerechtvaardigd. Er zijn waarnemingen gedaan waarbij zgn. soldaten als boodschappers optraden, die de werkers alarmeerden, en aan hen het vechten overlieten.  Fig. 68. Een termietenwerkster (links) en een termietensoldaat (rechts).
Ook in andere gevallen, bij andere dieren, waar geen ‘kasten’ onderscheiden worden, kan er toch tussen de leden van een gemeenschap verschil in bouw bestaan; dit geldt bijv. voor elke paringsgemeenschap. De verschillen bestaan hier soms alleen innerlijk, in de bouw der geslachtsorganen zelf; meestal echter hebben mannetje en vrouwtje ook verschillende hulporganen, zoals copulatie-organen en ev. ook signaalstructuren, die bij de balts dienst doen. De hierop berustende sexuele tweevormigheid kan in extreme gevallen zó ver gaan, dat men mannetje en vrouwtje onmogelijk als tot één soort behorend kan herkennen wanneer men niet toevallig weet dat zij uit ouders van één soort ontsproten zijn of als volwassen dieren met elkaar paren. Een extreem geval zijn de soorten waarbij zgn. dwergmannetjes voorkomen, zoals bij de in zee levende wormachtige Bonellia's. Een beschrijving | |
[pagina 103]
| |
hiervan vindt men in het boekje van Prof. Van Oordt over geslachtsverandering, dat als No. 4 van deze serie is verschenen. Veel talrijker echter zijn de gevallen waarin het verschil tussen de leden van een gemeenschap niet in de bouw tot uiting komt, maar uitsluitend uit het gedrag blijkt. Als voorbeeld kan ik weer de arbeidsverdeling bij de honingbij en wel speciaal die onder de werksters noemen. Men kan aan de bouw van een bij niet zien of men met een cellenpoetster of een honingdraagster te doen heeft; de verandering die zich in de bij voltrekt wanneer zij de ene functie tegen de andere verwisselt, uit zich slechts in het gedrag. Natuurlijk moet er, wil het gedrag veranderen, een of andere subtiele verandering binnen in de bij voltrokken zijn en wel met name in het zenuwstelsel, alleen men weet niet welke die verandering is. In één geval kent men de inwendige verandering wel, althans enigszins, nl. in het geval van de bouwbijen. Tegen dat een bij tot bouwster wordt, d.w.z. tegen dat zij aan de ratenbouw gaat deelnemen, ontwikkelt zich bij haar de zgn. wasklier, een huidklier aan het achterlijf, die de was naar buiten afscheidt. Ook hier echter moeten er nog fijne toestandsveranderingen in het zenuwstelsel plaats gehad hebben, om het mogelijk te maken, dat de wasafscheidende bijen nu ook met die was gaan bouwen. Uit dit alles blijkt wel, dat de onderscheiding: verschil in bouw - geen verschil in bouw, niet scherp is; er is altijd enig verschil, maar de ene keer blijkt dat duidelijk uit de bouw, de andere keer blijkt het verschil slechts uit het gedrag. Het is haast overbodig, andere voorbeelden van gedragsverschillen te noemen. Ik herinner aan enkele al eerder genoemde gevallen: één zilvermeeuw alarmeert, de andere reageren. Eén eend vliegt op, de hele troep volgt. Bij een bontbekplevierenpaar dat tegelijk jongen van het eerste broed en eieren van het tweede broed verzorgt, wisselen mannetje en vrouwtje hun gedrag voortdurend af tussen jongenleiden en broeden. Telkens is er dus één die broedt en één die jongen leidt, een verschil in gedrag dat niet afhankelijk is van verschil in sexe. | |
[pagina 104]
| |
En zo zou, indien men alle gevallen van samenwerking op de keper beschouwde, steeds blijken, dat er altijd verdeling van arbeid is. Zelfs bij de eenvoudigste vorm, die van het ‘meedoen’, is er steeds een ‘voorwerker’ en zijn er anderen, die, door het voorbeeld geprikkeld, meedoen. Ons volgende probleem is dus nu: hoe komt die diversiteit tot stand? Wat zijn de oorzaken die het verschil tussen de betrokken dieren doen ontstaan? Wij komen hiermee op een gebied, dat men gewoonlijk niet tot de sociologie rekent, hoewel het er logisch niet van te scheiden is. Men noemt de studie van de oorzaken der ontwikkeling meestal de ontwikkelingsmechanica. Enkele punten, die het dichtst bij de sociologie liggen zal ik heel kort behandelen. Kort vooral, omdat het onderzoek hier tot gespecialiseerd laboratoriumonderzoek geleid heeft, waaraan de veldwaarnemer (voor wie dit boekje in de eerste plaats bedoeld is), niets kan bijdragen; verder ook omdat in dit gebied ten dele door andere schrijvers in deze reeks al behandeld wordt. In het volgende wil ik dan ook niet meer dan een gedachtengang aangeven. De verschillen tussen de partners kunnen op de meest uiteenlopende wijze ontstaan, variërende van zeer subtiele en oppervlakkige inwerking tot zeer diep in het dier verankerde oorzaken. Er zijn er van zeer subtiele nerveuze aard, die, soms door uitwendige prikkels, plotseling optreden, zeer kort duren en even plotseling veranderen. Om een heel simpel geval te noemen: als in een troep vogels één eerder opvliegt dan de anderen en daardoor de hele troep ‘meesleurt’, is het verschil tussen actor en reactor (en) daarin gelegen, dat de eerste bijv. iets gevoeliger voor gevaar was dan de anderen. Soms zijn zulke nerveuze verschillen blijvend (juist als bij de mensen zijn er ‘bange’ en minder ‘bange’, oplettende en minder oplettende individuen), soms ook zijn zij tijdelijk, bijv. wanneer in een individu de schrik van een pas opgedane ervaring nog nawerkt. In vele gevallen, vooral van min of meer reciprook gedrag van twee partners of twee groepen van partners, | |
[pagina 105]
| |
zijn het de dieren zelf, die het gedrag door middel van zintuigelijke prikkeling bij elkaar induceren. Dat bijv. mannetje en wijfje van een zilvermeeuwenpaar, die beide over een broeddrift beschikken, maar toch niet beide tegelijk kunnen broeden, werkelijk om beurten broeden resp. rusten of voedselzoeken, ligt daaraan, dat de een de ander tot functieverandering kan stimuleren. Heeft één partner een tijdlang niet gebroed, dan welt de broeddrang langzaam aan weer in hem op. Hij gaat naar het nest, en uit zijn stemming door het uitvoeren van signaalbewegingen, die hij in verschillende graden van intensiteit tot zijn beschikking heeft. Meestal staat de partner al bij het eerste signaal op, zo niet dan volgt een intensiever signaal, en helpt dit nog niet, dan dringt de broedlustige vogel de ander eenvoudig van het nest. Hier staan de zaken dus zo: beide partners beschikken over dezelfde bewegingsmogelijkheden, de een induceert de ander tot het verrichten van de complementaire handelingen en omgekeerd. Op dergelijke wijze komt de arbeidsverdeling tussen man en vrouw bontbek tot stand; ook hier kan elk der partners beide rollen spelen en induceren ze bij elkaar de complementaire stemming. Bij de broedzorg van de Cichlide Hemichromus bimaculatus berust de arbeidsverdeling op een dergelijk mechanisme. Ook bij deze vis bewaken mannetje en vrouwtje om beurten de nestkuil met de eieren. De bewakende partner wordt, zoals door proeven is komen vast te staan, door het bezoek van de aflossende partner tot wegzwemmen geprikkeld. Zó sterk werkt het toezwemmen van de aflossende partner als signaal hiervoor, dat men wel heeft waargenomen, dat een wijfje van haar broed wegzwom als reactie op het naderbijkomen van een vreemde Hemichromus, aan wie zij zoodoende de eieren overliet. De vreemde echter, die niet, zoals het mannetje waarvoor het vrouwtje hem aanzag, broedlustig was, bleef gewoon rondzwemmen. Toen hij na enige minuten weer in de buurt van het vrouwtje kwam, vatte zij dit weer als een signaal op en nam haar plaats bij de eieren weer in. Dit alles ging van | |
[pagina 106]
| |
haar zijde gepaard met het bij een normale aflossing gebruikelijke ceremonieel. In vele gevallen van arbeidsverdeling echter liggen de verschillen tussen de partners stevig verankerd. Dit is al op het eerste gezicht duidelijk waar we met grote verschillen in bouw te doen hebben, zoals bijv. bij een bijenkoningin en een bijenwerkster, maar ook in gevallen waar zulke vormverschillen niet bestaan of niet zichtbaar zijn, kan een groot verschil in gedrag bestaan, dat niet alleen van buitenaf geïnduceerd wordt, zoals bijv. bij de winterkoning, waar mannetje en wijfje uiterlijk bijna niet verschillen, maar toch een heel verschillende taak in het huwelijk hebben. Deze en dergelijke verschillen komen tot stand door onomkeerbare ontwikkelingsprocessen, waardoor het ene individu zus, het andere zo groeit. Een paar voorbeelden volgen hier. In de eerste plaats kunnen de verschillen tussen de complementaire partners van een gemeenschap op leeftijdsverschil berusten. Elke jonge lijster spert, zolang hij nog in het nest verblijf houdt en niet al te verzadigd is, zodra de oude op het nest komt. Een erfelijk vastgelegd, onomkeerbaar ontwikkelingsproces heeft gemaakt, dat hij op dit ogenblik geen andere rol dan deze kân spelen. Hetzelfde geldt voor de moedervogel die op het sperren reageert door het jong te voeden. Er is hier natuurlijk geen kwestie van dat hier de rollen op een volgend ogenblik verwisseld kunnen worden. Leeftijdsverschil bepaalt verder ook de verschillen waarop de arbeidsverschillen tussen de werksterbijen berust. Op elk ogenblik is voor een werkster door leeftijd en erfelijke aanleg, en door de stand van haar groeiprocessen, bepaald welke rol zij zal spelen. Op erfelijk vastgelegde verschillen in ontwikkeling berusten ook bijna steeds de verschillen tussen de sexen. Veelal wordt reeds bij de bevruchting in de eicel vastgelegd tot welk geslacht het uit de eicel groeiende individu zal behoren. Weliswaar kan zulk een ontwikkeling bij sommige soorten door hormonen nog afgeleid worden; dit | |
[pagina 107]
| |
vergt dan echter steeds eveneens een lang ontwikkelingsproces, in tegenstelling met de bovenbesproken veranderingen van rol, die door prikkeling der zintuigen, langs nerveuze weg, geïnduceerd kunnen worden. Ik verwijs voor bizonderheden naar het boven aangehaalde boekje van Prof. Van Oordt. Alleen wil ik nog even wijzen op het feit, dat bij bepaalde Protisten de sexualiteit zgn. relatief is en dat daar de rol die een individu bij de paring speelt, geïnduceerd wordt door - nl. het complementaire gedrag is van - het gedrag van de tegenspelerGa naar eind87. Uit de verte vergelijkbaar hiermee is het abnormale gedrag dat vrouwelijke duiven en andere dieren vertonen, wanneer zij met andere wijfjes zonder mannetjes worden samengehouden; het vrouwelijk gedrag van een der dieren kan dan in een ander dier mannelijk gedrag inducerenGa naar eind88. Zulke nerveuze inducties zijn echter op sexueel gebied zeer zeldzaam, en meestal hebben wij dus werkelijk met erfelijk vastgelegde verschillen te maken. Weer een ander geval van door de ontwikkeling stevig vastgelegde verschillen tussen de partners van een gemeenschap zijn de verschillen tussen bijenkoningin en werksters. Beide ontstaan uit bevruchte, in aanleg gelijke eieren. Van hoeveelheid en kwaliteit van het voedsel dat de uit het ei gekomen larve krijgt hangt het echter af, of het ei zich tot werkster dan wel tot koningin zal ontwikkelen. De koninginnecellen zijn dan ook door hun grootte van de andere cellen in een broedraat te onderscheiden. Deze weinige voorbeelden zijn hopelijk voldoende, om de gedachtengang te illustreren. Het zal duidelijk zijn, dat er over deze kwesties heel wat meer te zeggen zou zijn; de verschijnselen, die hier slechts even werden aangeduid, behoren tot grote, tamelijk zelfstandige gebieden van wetenschap; vooral erfelijkheidsleer, ontwikkelingsphysiologie en celleer rekenen deze problemen tot hun domein. Daarom ga ik op deze kwesties niet verder meer in en sluit hierbij de behandeling van de analyse der sociale organisatievormen, om in de volgende hoofdstukken enige gedachtengangen van meer synthetische aard te bespreken. |
|