|
| |
| |
| |
6. De geheimen van één handeling
De stekelbaars kruipt door. Tijdens die doorkruip gebeurt er iets waardoor gedurende enige tijd het vermogen om nogmaals door te kruipen geblokkeerd wordt. De handeling zelf, kortom, voorkomt het optreden van een tweede soortgelijke handeling binnen korte tijd. De eerste vraag die rijst is: is de handeling inderdaad noodzakelijk om die blokkade te bewerkstelligen? Het zou best denkbaar zijn dat als T en E elkaar snijden, de T toch stijgt ook al vindt er geen doorkruipen plaats. De handeling doorkruipen zou best alleen maar een begeleidend verschijnsel kunnen zijn van de stijging van T zoals het verschijnen van rimpels op je voorhoofd een begeleidend verschijnsel kan zijn van denken, maar denken kan best plaatsvinden zonder dat het noodzakelijk is dat die rimpels verschijnen. Zodoende lag het voor de hand eerst te proberen of de stekelbaars de handeling misschien kan overslaan waarbij dan toch de blokkade tot stand komt. Daartoe lieten we, vlak voordat we een doorkruip verwachten, een glazen cilinder dalen over het nest van de stekelbaars zodat de stekelbaars zijn nest nog wel kon zien, maar er niet meer in kon doorkruipen op het moment dat dat te verwachten was. Steeds constateerden we daarbij dat de stekelbaars onmiddellijk na het weghalen van de cilinder, hoe lang die cilinder ook over het nest had gestaan, door het nest kroop. Met andere woorden: de doorkruip is inderdaad noodzakelijk, het dier moet de handeling uitvoeren. En pas als hij de handeling heeft uitgevoerd, begint het volgende interval want het tijdstip waarop de tweede doorkruip na het weghalen van de cilinder plaatsvindt is juist zoveel verschoven als het door de cilinder bewerkstelligde uitstel bedroeg.
Overslaan van doorkruipen is niet mogelijk. De handeling moet gebeuren. Kun je dan - dat is de volgende vraag - zodanig manipuleren met die handeling dat daaruit iets af te leiden valt over de T-factor? Om begrijpelijk te maken hoe ik kwam tot het volgende experiment moet ik een kleine zijsprong maken naar ons gedrags-genetisch onderzoek over de stekelbaars.
| |
| |
| |
Maniakken
Al in 1962 constateerde Sevenster dat er in het laboratorium opgekweekte mannetjes waren die na aanbieding van een vrouwtje niet eenmaal, maar tweemaal achter elkaar doorkropen. Met deze mannetjes kweekte Sevenster verder en ook in de volgende generaties werd gevonden dat dieren van deze laboratoriumstam tweemaal, ja soms drie-, vier-, of vijfmaal doorkropen binnen twee minuten. Zo af en toe is er zelfs een visje dat nog vaker door zijn nest gaat. Al deze herhaaldelijk doorkruipende dieren werden maniakken genoemd.
Vaker dan eenmaal doorkruipen binnen twee minuten past niet in het model van Nelson, is daar zelfs strijdig mee want in het model wordt voorondersteld dat de handeling doorkruipen nogmaals optreden van die handeling gedurende enige tijd blokkeert. Alleen al daarom leek het gewenst ze nader te onderzoeken. Uit onderzoek van Sevenster en mij is gebleken dat dit zogenaamde dubbel doorkruipen waarschijnlijk een erfelijk vastgelegde gedragsafwijking is. Ik zal op dit onderzoek, hoe interessant ook, niet nader ingaan. Slechts dit is nu voor ons van belang: bij bewerking van de door Sevenster en mij bijeengebrachte gegevens over het dubbel-doorkruipen bleek dat er een grotere kans is op tweemaal doorkruipen naarmate de eerste doorkruip van zo'n serie van twee (of meer) korter duurt. Daaruit vloeide onmiddellijk de vraag voort: kun je de kans op tweemaal doorkruipen vergroten als je het nest van het dier verkort? Indien je het nest verkort, verkort je ook de nesttunnel en is de door de stekelbaars af te leggen weg tijdens doorkruipen kleiner. Dan moet ook de doorkruipduur korter zijn tenzij de stekelbaars plotseling langzamer zou gaan doorkruipen. Inderdaad bleek dat je de kans op het optreden van dubbel-doorkruipen kon vergroten door nest-verkorting uit te voeren. Het lag voor de hand nu ook te gaan proberen of je van een normaal dier dat nooit dubbel doorkroop een maniak kon maken door zijn nest te verkorten. Daarom verkortte ik nesten van normale dieren. Je moet daarbij met een schaar een stuk van het nest afknippen en wel zodanig dat het overblijvende deel noch beschadigd noch verschoven wordt. Het is vooral van belang dat de nestopening op zijn plaats blijft; als die opening ook maar iets verschuift zal de stekelbaars dat nest verder niet meer accepteren doch elders een nieuw nest gaan bouwen.
| |
| |
| |
Het nest verkort
Als ik van normale dieren het nest verkortte, volgde geen dubbel doorkruipen. In plaats daarvan constateerde ik tot mijn grote verbazing dat de eerstvolgende doorkruip, na een doorkruip door een verkort nest, eerder kwam dan ik verwachtte. Bij één van de eerste dieren, waarbij ik het nest verkortte, had ik na een doorkruipduur van 7 sec altijd een doorkruipinterval gemeten van bijna 21 minuten. Nu mat ik na een doorkruipduur van 4 seconden een doorkruipinterval van iets meer dan 12 minuten. Het grappige is dat de stekelbaars zijn nest na een verkorting na verloop van tijd weer aanbouwt zodat je het ook een tweede keer kunt verkorten. Je kunt uiteraard op verschillende plaatsen een stuk van het nest afknippen, of vlak na de opening, of halverwege, of vlak voor het uiteinde van het nest. In al die gevallen zul je een verschillende doorkruipduur meten bij verschillende nestlengten. Hoe korter het nest, hoe korter de duur van het doorkruipen. Het verrassende daarbij is dat bij elke duur van doorkruipen een passende intervallengte behoort en wel zodanig dat er een recht evenredig verband is tussen duur van doorkruipen en daaropvolgend interval. Je kunt zoiets wel enigszins begrijpen naar analogie van slapen bij mensen. Ben je gewend normaal 8 uur per nacht te slapen dan zul je na een nacht van 4 uur slaap eerder slaperig zijn dan anders en na een nacht van 2 uur slaap zul je nog eerder weer naar bed verlangen. Of vergelijk het met eten: eet je bij het ontbijt half zoveel als gewoonlijk dan zul je veel eerder trek krijgen dan anders. Neem je alleen een kop thee met een beschuit dan ben je na een uur al weer hongerig. Zoiets moet er ook aan de hand zijn bij de stekelbaars, maar het vreemde is dat het zo akelig precies in elkaar steekt. Het lijkt wel alsof dit visje een ingebouwde chronometer heeft!
Je kunt ook nog op een andere manier de duur van het doorkruipen verkorten. Als de stekelbaars bezig is met doorkruipen, kun je naar het aquarium, waarin het zich afspeelt, toesluipen en de stekelbaars met een lange glazen staaf een tik op de staart geven. Zodra je dat doet, zal de stekelbaars onmiddellijk zijn nest verlaten. Je kunt zo'n tik op verschillende tijdstippen tijdens doorkruipen toedienen en op die manier een aantal verschillende, te korte doorkruipduren verzamelen en de daarna komende intervallen meten. Ook dan blijkt er een recht evenredig verband te zijn tussen de doorkruipduur en de lengte van het daaropvolgende doorkruipinterval. Uit dit alles kun je concluderen
| |
| |
dat de duur van de doorkruiphandeling van groot belang is.
| |
Open dak
Er is ook nog een andere mogelijkheid om de duur van doorkruipen te verkorten. Je kunt het nestdak overlangs openknippen. Daartoe steek je een punt van de schaar voorzichtig in de nesttunnel en knip je vervolgens het dak in tweeën. Stekelbaarzen kruipen sneller door in nesten met open daken, waarschijnlijk omdat de te overwinnen weerstand van het nest tijdens doorkruipen geringer is. Je kunt de doorkruipbeweging dan bovendien beter waarnemen. Ook in dat geval vind je dat er een recht evenredig verband is tussen de doorkruipduur en de lengte van het daarna komende doorkruipinterval. Uit dit experiment kun je tevens opmaken dat de druk van zijwanden en bovenkant van het nest, die een door de stekelbaars tijdens doorkruipen te registreren prikkel zou kunnen zijn die de T-factor doet stijgen, blijkbaar niet zo belangrijk is. Als het dak open is, is de mechanische druk van het nest heel anders dan normaal. Toch vind je normaal, zij het korter dan anders durend doorkruipen en ook nu is slechts die duur van doorslaggevende betekenis. Uit dit experiment valt ook op te maken dat het in de duisternis van de tunnel verkeren niet de tijdens doorkruipen te registreren prikkel kan zijn die de T-factor doet stijgen want als het dak open is, blijft de doorkruipende stekelbaars licht zien.
Als er niets met het nest gebeurd is, blijft de doorkruipduur constant. Maar een heel enkele stekelbaars heeft van nature al een variërende doorkruipduur ook al doe je niets met het nest. In één geval had ik een visje dat een van 10 tot 22 seconden variërende doorkruipduur had en daarmee in overeenstemming was dat hij ook een variërend intervallengte had, waarbij elk interval bleek te passen bij de daaraan voorafgaande doorkruipduur.
| |
Nest verlengen
Vanzelfsprekend rijst nu de volgende vraag: wat gebeurt er als de duur van het doorkruipen langer is dan normaal? Is het daarna komende dk-interval dan ook recht evenredig veel langer of niet? Experimenteel bleek deze vraag moeilijk te benaderen te zijn. Om lange doorkruipduren te verkrij- | |
| |
gen moest de lengte van de nesttunnel langer zijn dan normaal. Hoe dat te bereiken? Ik probeerde eerst of ik een tweede nest (van een andere man) achter het nest van mijn proefdieren zodanig kon laten aansluiten dat ik een langer nest verkreeg. Je kunt een nest gemakkelijk oppakken, het is zo stevig gebouwd dat het niet uit elkaar valt. Ik knipte het uiterste einde van het nest van mijn proefdier af, knipte het begin van een elders geroofd tweede nest af en legde de beide nesttunnels op het snijvlak tegen elkaar aan. In de meeste gevallen komt de stekelbaars je, na afloop van het tegen elkaar aan leggen van beide nesten, te hulp. Hij stampt op het nest, plakt over het snijvlak heen en kit zo zelf beide nesten aan elkaar. Tot zover gaat het dan ook vaak goed. Maar als je vervolgens een experiment gaat uitvoeren (vrouwtje erbij, waarnemen van doorkruipintervallen na doorkruipen door een verlengd nest) wacht je haast altijd een grote teleurstelling. De stekelbaars kruipt wel door in het verlengde nest, maar kruipt er doorgaans niet helemaal doorheen doch komt door dak of zijwand naar buiten zodra hij zijn normale duur van doorkruipen heeft volgemaakt. Het lijkt wel of hij dan ‘weet’: ik moet er nu uit, ik heb lang genoeg doorgekropen want slechts bij hoge uitzondering kruipt zo'n stekelbaars langer door dan normaal. Op zich is dit wel een leuk resultaat, je kunt er misschien uit concluderen dat er blijkbaar een soort ‘Sollwert’ voor doorkruipduur bestaat. Toch is het wel gelukt dieren langer dan normaal te laten doorkruipen. Ik heb er echter zeer veel nesten voor moeten verlengen. In alle gevallen waarin de doorkruip langer duurde dan
normaal, duurde het daarna komende interval niet langer dan normaal. Na verlengde doorkruipen vind je normale intervallen. Eén dier kwam mij bij dit experiment op bijzondere wijze te hulp. Ik deed met dit dier nestverkortingsexperimenten en knipte daarom het nest in tweeën. Het
achterste gedeelte van het nest legde ik een eindje bij het voorste gedeelte vandaan zoals ik dat altijd deed. Dit dier sloopte dat afgeknipte gedeelte niet zoals de andere dieren altijd deden, maar bouwde een stukje nest tussen de twee losse delen van het nest in zodat een langer nest ontstond. In dat langere nest kroop hij ook langer door, maar de lengte van zijn doorkruipinterval was daarna niet langer dan normaal. Weer knipte ik het verlengde nest in tweeën, weer legde ik het achterste stuk een eindje weg en opnieuw bouwde deze stekelbaars er een stukje nest tussen in. Zo verkreeg ik na vijf keer knippen en vijf keer herbouw een vrij lang nest waarin het dier ook vrij
| |
| |
lang doorkroop, veel langer dan normaal in ieder geval. Maar nooit heb ik een verlengde interval gevonden. Na vijf keer herbouw gaf hij het trouwens op, hij bouwde elders een nieuw nest van normale lengte.
Er is ook nog een andere mogelijkheid om de duur van het doorkruipen te verlengen. In plaats van het nest te verlengen, kun je het dier ook, als hij op het punt staat om het nest te verlaten na een doorkruip, in het nest vasthouden. Je laat dan een glazen staaf met een verbreed uiteinde voorzichtig op het nest zakken. Het dier kan nu het nest niet verlaten, maar zal het wel proberen; hij slaat, net als bij het gewone doorkruipen, voortdurend met zijn staart. Je kunt het dier op deze wijze elk gewenst aantal seconden in het nest vasthouden en zo behoorlijk lange duren van doorkruipen induceren. Ook dan bleek dat er daarna nooit verlengde intervallen gevonden werden. Toch zegt de uitslag van dit experiment minder dan de uitslag van het verlengde-nestexperiment. Het zou namelijk best zo kunnen zijn dat een stekelbaars die je in zijn nest vasthoudt helemaal niet meer aan het doorkruipen is, doch alleen nog maar wanhopig probeert om uit zijn nest te komen. Die wanhopige pogingen om het nest te verlaten lijken wel veel op doorkruipen, maar zijn het misschien niet. Om nu vast te stellen of dat inderdaad het geval is kun je het volgende experiment doen. Je laat de stekelbaars in zijn nest kruipen en je begeeft je daarna met de glazen staaf naar het nest. Je geeft hem een tik op zijn staart en tegelijkertijd laat je de glazen staaf met het verbrede uiteinde op het nest zakken zodat het dier het nest niet meer uit kan. Ook dan zul je zien dat de stekelbaars, slaande met zijn staart, uit het nest probeert te komen en al zijn bewegingen daarbij lijken vrij veel op het normale doorkruipen. Als dit gedrag inderdaad als normaal doorkruipen geldt, moet je vinden dat de stekelbaars daarna opnieuw in zijn nest zal doorkruipen na een tijd die in overeenstemming is met de totale duur van het doorkruipen tot staaf op nest plus het dier vasthouden in zijn nest tot een maximum van de normale doorkruipduur. Als die pogingen om uit het nest te komen nadat hij de glazen staaf op zijn
nest krijgt niet gelden als normale doorkruiphandelingen, moet je vinden dat het dier daarna weer zal doorkruipen tot het moment waarop je hem een tik gaf met de staaf. Inderdaad blijkt altijd het laatste het geval te zijn: de stekelbaars probeert wel uit zijn nest te komen en dat proberen lijkt wel op doorkruipen, maar ‘telt’ niet als doorkruipen zoals je aan de lengte van het daarna komende interval kunt aflezen. De
| |
| |
op doorkruipen lijkende bewegingspatronen zijn niet voldoende om de T-factor te doen stijgen, het dier moet ook in een ‘doorkruipstemming’ zijn om nu even een gevaarlijk woord te gebruiken. Is het dier dat niet doordat je hem een tik op de staart hebt gegeven en vasthoudt in zijn nest, dan is er ook geen sprake van echt doorkruipen meer. Ook dat wijst er op dat datgene wat zich in het dier afspeelt kennelijk heel belangrijk is.
| |
Is het wel de T-factor?
In het voorgaande heb ik steeds, impliciet, verondersteld dat het verkorten van het nest inderdaad invloed heeft op de T-factor. Is dat echter wel juist? Het is toch best denkbaar dat zo'n nestverkorting de E-factor beïnvloedt. Als manipuleren met het nest inderdaad E beïnvloedt dan moet, volgens het model van Nelson, de E-factor onder invloed van nestverkorting stijgen. Alleen dan kun je een verkorting van de lengte van het doorkruipinterval verwachten. We kunnen nu het volgende experiment doen om na te gaan of er invloed is op de E-factor als we het nest verkorten. We nemen drie achtereenvolgende intervallen van doorkruipen waar (zie nogmaals afb. 46). Op tijdstip A nemen we een normale doorkruip waar, op tijdstip B evenzo. Direct na de doorkruip op tijdstip B verkorten we nu het nest. In het interval BC heeft het visje dus steeds een verkort nest, maar dat interval is begonnen met een doorkruip van normale duur. Op tijdstip C vindt een doorkruip plaats in het verkorte nest en zal dus ook de duur van het doorkruipen verkort zijn. Als er nu een directe invloed is van het verkorten van het nest op de E-factor moet ook het interval BC al verkort zijn; als er daarentegen slechts indirect via de duur van het doorkruipen invloed is op de lengte van het interval zal alleen het interval CD maar verkort zijn. In werkelijkheid vinden we dat ook. Er is nooit gevonden dat interval BC korter (of langer) is dan normaal. Ondanks het feit dat we in dat interval het nest van het visje verkorten vinden we een interval van normale lengte. Overigens rijst bij menige lezer misschien de vraag of de stekelbaars dat nu allemaal maar accepteert, al dat geknoei met zijn nest, een grote hand in zijn aquarium waarin bovendien een schaar hangt. Het antwoord daarop moet luiden dat de stekelbaars alleen dan al zulke ingrepen tolereert als hij heel tam is. In tegenstelling tot tiendoornige stekelbaarzen die om de een of andere
| |
| |
reden zelden schuw worden, zijn driedoorns betrekkelijk gauw schuw. Hoe dat precies komt, weten we nog steeds niet. Wel weten we dat sommige mensen stekelbaarzen uiterst tam kunnen maken (ik behoor om voor mijzelf onverklaarbare redenen ook tot die mensen, terwijl ik toch helemaal niet zo rustig ben) en dat andere mensen, hoe ze ook hun best doen, stekelbaarzen al na een paar uur voor altijd diep achter in het aquarium onder een hoopje flap weten te jagen. Zulke mensen zijn in het algemeen hoekige, voorzichtige bewegers. Mensen met een aarzeling in hun manier van doen of, wat ook kan, mensen die erg nerveus van aard zijn.
Stekelbaarzen tolereren deze ingrepen slechts mits ze tam zijn. Als ze heel tam zijn, gebeurt het vaak dat ze, terwijl je bezig bent het nest voorzichtig stuk te knippen, in je vinger proberen te bijten of tegen je schaar aanduwen. Ze zien, met andere woorden, wat er gebeurt. Toch heeft dat zien geen meetbare invloed op het doorkruipinterval.
Ook op een andere wijze kunnen we aannemelijk maken dat de nestverkorting geen invloed heeft op de E-factor. Als er invloed zou zijn van nestverkorting, zou die moeten bestaan in het verhogen van het niveau van de E-factor. Daar, zoals we al eerder zagen, zigzaggen te omschrijven is als de som van E plus T zou na nestverkorting E plus T groter moeten zijn. Met andere woorden: er zou na nestverkorting meer gezigzagd moeten worden. In werkelijkheid is dat niet het geval en daarom is het uitermate onwaarschijnlijk dat E rechtstreeks beïnvloed wordt door de ingreep nestverkorting.
Het is interessant na te gaan wat er gebeurt met het gedrag van de stekelbaars in het verkorte interval. Wat doet het waaieren, wat doet het boren? In een gewoon doorkruipinterval vind je na doorkruipen weinig of geen waaieren en naarmate de tijd verstrijkt zie je dat het waaieren toeneemt. In het verkorte interval vind je bijna altijd dadelijk na de doorkruip in het verkorte nest al weer waaieren en hoe korter de doorkruip duurt, des te meer waaieren zie je meteen na het doorkruipen al optreden. Het lijkt erop dat dat eerste deel van het interval gewoon wordt overgeslagen als het nest verkort is en als gevolg daarvan ook de doorkruipduur verkort is. Je kunt zelfs heel precies mathematisch analyseren hoe het verloop van het waaieren is in normale en verkorte intervallen en dan blijkt dat een stekelbaars in een verkort interval juist zoveel waaiert als overeenkomt met de hoeveelheid in een normaal interval in
| |
| |
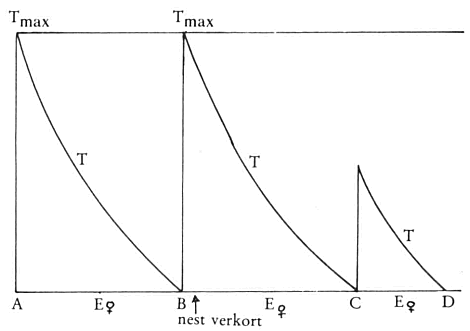
Afb. 51. Wat T doet als het nest verkort is
dezelfde minuut, teruggerekend vanaf het tijdstip waarop de volgende doorkruip het verkorte interval beëindigt. Als het normale interval van een stekelbaars 30 minuten duurt en het verkorte bijvoorbeeld 8 dan lijken die 8 minuten, wat het gedrag betreft, precies op de laatste 8 minuten van het normale interval. De eerste 22 minuten zijn gewoon ‘overgeslagen’. Ook het boren, normaal pas vlak voor een volgende doorkruip optredend, treedt in een verkort interval veel eerder op en ook dan correspondeert de hoeveelheid boren met de normaal te vinden hoeveelheid in het overeenkomstige laatste gedeelte van het normale interval.
Is dit nu te begrijpen in termen van het model van Nelson? Ja, heel goed. Waaieren is, zoals we zagen, uit te drukken als het verschil tussen E en T. Dat de stekelbaars in een verkort interval al zoveel meer waaiert duidt erop dat T lager is dan normaal (E blijft immers gelijk). Als we nu simpelweg aannemen dat die T-factor onder invloed van doorkruipen stijgt tot Tmax in een normale situatie, maar onder invloed van een verkorte doorkruip juist zoveel stijgt als in overeenstemming is met de duur van het doorkruipen dan zijn alle resultaten van onze waarnemingen daarmee te verklaren in termen van het model. Kijk maar naar afb. 51. T stijgt minder hoog en daarom lijkt het daarna komende interval precies op het laatste deel van een gewoon interval want T daalt in dat laatste gedeelte op dezelfde wijze.
| |
| |
| |
De compensatie
Zoals we al eerder zagen, is er blijkbaar een soort ‘Sollwert’ voor de doorkruipduur. Een stekelbaars wil niet langer in zijn nest verblijven dan zijn normale doorkruipduur duurt. Dan is T gestegen tot Tmax en hoeft hij ook niet langer te blijven liggen, zou je in termen van het model gesproken, kunnen zeggen. Dat er inderdaad zo'n soort Sollwert is, een streefwaarde, blijkt ook uit iets heel anders. Als je een nest verkort zal een driedoorn het na verloop van tijd weer aanbouwen. Sommige stekelbaarzen wachten heel lang met aanbouwen, andere beginnen vrij snel na het doorkruipen al materiaal aan te dragen en te bouwplakken. Als een stekelbaars vrij lang wacht met herbouw zal hij niet één keer, maar een aantal malen achter elkaar door een verkort nest heen kruipen. Het gekke daarbij is dat hij elke volgende keer dat hij door het verkorte nest heen gaat, langer over de doorkruip doet ook al is het nest nog helemaal niet aangebouwd. Dat houdt in dat hij op de een of andere manier de af te leggen weg langzamer ‘neemt’. Je kunt dat ook wel zien, hij blijft gewoon wat langer in het nest liggen. Hij compenseert voor nestverkorting met een langer verblijf in het nest. En dat blijkt ook zin te hebben want na zo'n langere doorkruip in een verkort nest is ook het daarna komende interval langer en wel precies zoveel langer als je op grond van de gemeten doorkruipduur verwacht. Ook dat wijst erop dat er een soort streefwaarde is voor de duur van doorkruipen. De stekelbaars bemerkt blijkbaar dat hij te kort doet over zijn doorkruip en blijft daarom langer liggen. Misschien is dat ook wel de reden waarom hij het nest aanbouwt, hij heeft een nest nodig waarin hij een normale duur van doorkruipen kan maken.
| |
Wat gebeurt er nu tijdens doorkruipen?
Nu we eenmaal weten dat de T-factor onder invloed van de duur van het doorkruipen juist zoveel stijgt als het doorkruipen aan tijd in beslag neemt, is het mogelijk dat gegeven te gebruiken voor verdere experimenten. Op de één of andere wijze moet de stekelbaars precies kunnen registreren hoe lang hij over zijn doorkruip doet want anders zou het niet mogelijk zijn dat die duur zo'n grote invloed zou kunnen hebben. Hoe registreert de stekelbaars die duur van doorkruipen nu zo precies? Is er één of andere uitwendige
| |
| |
prikkel die hij met zijn zintuigen waarneemt welke voor de precieze registratie van de duur zorgt? Is het registratie van de uitvoering van de handeling, met andere woorden, registratie van zijn spierbewegingen met behulp van zenuwuiteinden in de spieren. Of wordt het geheel intern geregistreerd? In het laatste geval zouden we, als we meer willen weten, in het dier moeten ingrijpen, hersen-laesies moeten aanbrengen of zenuwen moeten doorsnijden, of hormoonklieren moeten weghalen.
Als het simpelweg registratie van een uitwendige prikkel is, moeten we in staat zijn een kunstmatige uitwendige prikkel aan te bieden ter vervanging van de stimulatie tijdens doorkruipen, overeenkomstig de wijze waarop bij een daarvoor ontvankelijke man een mooie foto van een knap meisje al prikkelend werkt. Sevenster bedacht daarom het volgende experiment. Op het moment dat wij doorkruipen verwachten, laten we de stekelbaars door een tunneltje kruipen dat op de nesttunnel lijkt. Als ook dat een stijging van de T-factor veroorzaakt, zijn we een stap verder. Maar het is uiteraard niet zo gemakkelijk om een stekelbaars door een tunneltje van jouw keuze te laten kruipen. Sevenster plaatste nu een schot in een aquarium waarin een driedoornstekelbaars zijn nest had gebouwd en in dat schot werd een trechter aangebracht in welks uiteinde schuimplastic werd gedaan. Achter het schot kun je een rijp vrouwtje aanbieden in een buis en als je dat trechtertje nu vastmaakt in een op te halen luik, kun je, als je het luik opendoet, de stekelbaars gemakkelijk achter het schot lokken. Zodra hij erachter is en baltst tegen het rijpe vrouwtje in de buis, kun je het schot laten zakken zodat de stekelbaars door trechter en trechteruiteinde met schuimrubber terug moet naar het nest. Je moet dat een stekelbaars eerst wel even leren, maar de neiging om terug te gaan naar het nest is zo sterk dat je daar goed gebruik van kunt maken. Zodoende kun je de trechter inderdaad alleen dan laten zakken als je een doorkruip verwacht en daarmee de stekelbaars een stimulus-substituut voor het nest aanbieden. Noch uit de experimenten van Sevenster, noch uit die van mijzelf met de trechter is ooit gebleken dat het door de trechter gaan doorkruipen kan vervangen, zelfs niet als je in de trechter in plaats van schuimrubber groene alg aanbrengt. Met andere woorden: een op de natuurlijke stimulus gelijkende prikkel, aangeboden op het moment van een te verwachten doorkruip, heeft geen met het nest
vergelijkbare werking. Helaas.
Daarna hebben we het volgende gedaan. Moet het dier altijd
| |
| |
door zijn eigen nest heenkruipen? Kan alleen het eigen nest de registreerbare prikkel leveren? Om dat te onderzoeken was een nogal vernuftige proefopzet noodzakelijk die ik niet zal beschrijven, maar die het resultaat opleverde dat de stekelbaars niet door zijn eigen nest hoeft te gaan om de T-verhoging te bewerkstelligen. Ook als hij op het moment van een te verwachten doorkruip door een vreemd nest gaat, stijgt de T-factor, dat wil zeggen, wordt er lange tijd niet doorgekropen. Het verschil met de trechter met schuimrubber is dat de stekelbaars nu een nest heeft en zelf daarin doorkruipt terwijl we hem bij de trechter dwongen om door die trechter heen te gaan.
| |
Wordt er iets uitgescheiden of juist aangemaakt
tijdens doorkruipen?
Je zou je kunnen voorstellen dat er tijdens het doorkruipen iets geleegd wordt dat daarna weer wordt aangemaakt. Kruipt de stekelbaars door een verkort nest dan krijgt hij maar een bepaald gedeelte van de normale tijd gelegenheid iets te legen dat daardoor eerder weer gevuld is zodat eerder dan anders opnieuw doorkruipen zal plaatsvinden. Het kan ook dat hij iets aanmaakt tijdens doorkruipen en daarvan te weinig aanmaakt tijdens een verkorte doorkruip. Maar in geval van aanmaken van iets moet dat aangemaakte dan tijdens het volgende doorkruipinterval verdwijnen zodat, als het op is, de volgende doorkruip volgt.
Het is razend moeilijk deze veronderstellingen te onderzoeken. Als hij iets leegt tijdens doorkruipen zou je dat chemisch moeten kunnen aantonen. Hij kan niet de spermablaas legen, zoals we al eerder zagen. Hij zou bijvoorbeeld ook een zekere hoeveelheid plaksel kunnen afscheiden tijdens doorkruipen. Als je echter het nest openmaakt na een doorkruip zie je geen plakdraad. Dat kan het dus ook niet zijn. Urine dan? Onwaarschijnlijk. Bovendien zitten de van de nieren aflopende leiders vol met plaksel.
Als hij tijdens doorkruipen iets aanmaakt (bijvoorbeeld een hormoon) dat daarna weer wordt afgebroken in het lichaam, zou je ook alleen door ingrepen in het lichaam daarover iets te weten kunnen komen.
Je zou ook nog kunnen veronderstellen dat de toestand van het nest doorkruipen veroorzaakt. In het nest zou een stof, bijvoorbeeld koolzuur (het nest bestaat immers uit nog levende plantedelen) kunnen worden opgehoopt. Is er een
| |
| |
maximum aan koolzuur dan kruipt hij door. Maar ook dat is onwaarschijnlijk aangezien de stekelbaars niet onmiddellijk doorkruipt in een lang niet gebruikt nest en zo'n nest kun je hem aanbieden met behulp van de eerder beschreven draaitafel. In het nest zou ook een stof kunnen verdwijnen en als deze op is vindt de volgende doorkruip plaats. Maar ook dat is uit te sluiten op grond van het experiment met de draaitafel. Ook dan zou een lang niet gebruikt nest na aanbieding onmiddellijk doorkruipen oproepen.
Behoudens de mogelijkheid dat hij in het lichaam tijdens het doorkruipen iets aanmaakt of juist afbreekt dat van buiten niet of nauwelijks meetbaar is, moeten we alle mogelijkheden dat er stoffen aangemaakt of juist afgebroken worden tijdens doorkruipen verwerpen. Wat dan? Eerder had ik het al over een periode van duisternis in de tunnel.
| |
De donkerperiode in de tunnel
Percipieert de stekelbaars met zijn ogen de duur van de duisternis (of althans verminderde lichthoeveelheid) tijdens het doorkruipen? Dat vast en zeker, maar is dat dan ook de effectieve prikkel? Alleen al op grond van het feit dat een dier dat door een nest met open dak doorkruipt, daarmee toch een doorkruipinterval laat registreren, kunnen we verwerpen dat het om lichtprikkels zou gaan. Maar we treden in ons laboratorium niet spoedig over één nacht ijs. Je kunt de stekelbaars ook een nest laten bouwen van doorzichtige plastic nylon draadjes en ook dan zal hij gewoon doorkruipen. Daarna is niettemin sprake van een interval. Als wij vlak voordat wij een doorkruip verwachten even het licht uitdoen en vervolgens weer aan zodat de stekelbaars op het moment dat T en E elkaar snijden in de duisternis zit, maar niet in het nest, blijkt dat het dier na het aangaan van het licht alsnog doorkruipt. Nee, het is niet waarschijnlijk dat licht/donker de werkzame prikkel is.
| |
De druk van het nest
Wat dan? Kan het zijn dat de stekelbaars de druk die het nest uitoefent registreert met zenuwuiteinden in de huid of met het zijlijnsysteem? Vast en zeker. Maar is dat de werkzame prikkel die de T-factor doet stijgen? Niet waarschijnlijk omdat dieren met open nestdaken normaal doorkropen
| |
| |
terwijl je toch moet aannemen dat zo'n open dak mede tot gevolg heeft dat het nest een minder krachtige druk uitoefent op het lichaam van het dier. We kunnen ook langs andere weg experimenteel bevestigen dat het waarschijnlijk niet de mechanische druk is die het hem doet. Als we met behulp van elastiek over het nest heen de druk van het nest kunstmatig vergroten zien we dat de stekelbaars toch door het nest heenkruipt al kost het hem zichtbaar meer moeite. Daar T echter nooit hoger lijkt te stijgen dan Tmax hebben we er niet veel aan om op deze manier de druk van het nest te vergroten want we zullen, ook al zou die extra druk geregistreerd zijn, toch nooit een langer interval dan normaal waarnemen. Inderdaad blijkt dat ook het geval te zijn. Maar als je nu een stekelbaars door een met elastiek verzwaard nest laat doorkruipen en nog tijdens de doorkruip een tik op de staart geeft zodat hij het nest eerder dan gewoonlijk verlaat, heeft hij gedurende de tijd van doorkruipen in het nest een grotere druk dan normaal ervaren. Als zoiets effect heeft, moet dat in de duur van het volgende interval zijn af te lezen. Maar hoeveel experimenten van dit type ik ook gedaan heb, nooit heb ik enig effect van de vergrote druk kunnen merken. Ook dat, zo lijkt het, kan niet de werkzame prikkel zijn, al kun je hiermee niet uitsluiten dat de gewone druk niet de effectieve prikkel zou kunnen zijn. Maar op grond van de resultaten in nesten met open daken valt ook dat als mogelijkheid eigenlijk weg.
| |
Staartslaan
Is het dan denkbaar dat de stekelbaars zijn eigen spierbewegingen registreert en dat deze registratie de werkzame prikkel levert die T doet stijgen? Tijdens het doorkruipen slaat de stekelbaars met zijn staart en dat staartslaan zou natuurlijk best eens de werkzame prikkel kunnen zijn. Het zou dan net zoiets zijn als de slagen waarmee je je horloge opwindt; elke volgende slag stelt het moment waarop je opnieuw moet opwinden weer enige tijd uit.
Als het het staartslaan is dat het hem doet, moet er een precieze correlatie zijn tussen de duur van het doorkruipen (die immers zo belangrijk is) en het staartslaan. In werkelijkheid blijkt die correlatie geenszins aanwezig te zijn, een dier dat altijd precies 15 seconden doet over zijn doorkruip kan twaalf en de volgende keer bijvoorbeeld vijftien staartslagen produceren. Op grond van wiskundige berekenin- | |
| |
gen moet het helemaal uitgesloten worden geacht dat het inderdaad het staartslaan is dat de T-factor doet stijgen. Er zijn, los daarvan, nog een paar argumenten voor. Als de stekelbaars compenseert voor nestverkorting met langer in het nest blijven liggen, slaat hij niet vaker met zijn staart dan wanneer hij heel kort doorkruipt in het verkorte nest, maar blijft hij inderdaad langer liggen. De tussenpozen tussen staartslaan zijn dan groter. Ook dat wijst erop dat het niet het staartslaan is dat T doet stijgen. Voorts kun je tijdens doorkruipen naar het aquarium waarin dat gebeurt toelopen en een tik op de ruit geven. Sommige stekelbaarzen willen dan wel eens blijven liggen in het nest en tegelijkertijd ophouden met staartslaan. Hoewel de duur van het doorkruipen dan niet afwijkt van de normale duur, is het aantal staartslagen minder dan anders. Toch is daarna ook een normaal doorkruipinterval meetbaar. Dit lukt overigens slechts heel zelden, maar wijst er toch ook op dat het staartslaan als werkzame factor uitgeschakeld kan worden.
| |
De onderkant van het lichaam
Doorkruipen begint altijd met boren en bij het boren drukt de stekelbaars zijn keel tegen de ondergrond van het nest. Als de stekelbaars het nest verlaat na doorkruipen, is het gedeelte van het lichaam vóór de anale vin het laatst in contact met de ondergrond. Als het dak van het nest openligt is toch altijd nog de onderzijde van het lichaam in contact met het nest en wordt die onderzijde geprikkeld door de beweging van het doorkruipen. Zitten er dan misschien in de onderzijde van het lichaam huidzintuigjes die de duur van het doorkruipen registreren? Om dit te onderzoeken leek het gewenst de bodem van het nest te veranderen. Daarom liet ik heel dunne en iets ronde glasplaatjes slijpen in de beroemde glasblazerij van het Fysisch Laboratorium in Leiden en legde ik deze glasplaatjes in nesten van stekelbaarzen (nadat ik eerst het dak had opengemaakt). De eerste stekelbaars waarbij ik dat deed gleed tijdens de doorkruip over het plaatje, keerde meteen terug naar de nestopening en probeerde opnieuw door te kruipen, gleed weer uit over het plaatje en probeerde een derde doorkruip. Op dat moment dacht ik verheugd: nu heb ik het gevonden, maar helaas, dat bleek toch niet het geval te zijn. Alle stekelbaarsjes die geconfronteerd worden met een glasplaatje op de bodem van het nest glijden aanvankelijk uit en
| |
| |
proberen daarom nogmaals door te kruipen, maar op de lange duur leren zij toch om de juiste bewegingen te maken over het glasplaatje en dan kruipen ze door alsof er niets aan de hand is en vind je na een doorkruip over zo'n glasplaatje ook een normaal doorkruipinterval. Wellicht is er inderdaad sprake van dat het dier met de onderzijde van het lichaam iets registreert, maar strikt nodig is het blijkbaar niet want ook op een volkomen glad oppervlak dat toch een heel andere prikkel aflevert dan een ruwe nestbodem, is een normale doorkruip uit te voeren. Het gaat er maar om dat de stekelbaars de juiste beweging maakt.
Niet eens zo heel lang na de experimenten met glasplaatjes bezat ik een stekelbaars die wel in seksuele fase was (dat wil zeggen wel baltste tegen een vrouwtje dat in zijn aquarium werd geplaatst) maar niet beschikte over een nest. Hij had ooit wel een nest gehad, maar dat afgebroken en geen nieuw gebouwd. Als je deze stekelbaars een goed baltsend vrouwtje aanbood, ging hij naar een bepaald plekje op het zand in het aquarium (niet de plaats waar hij vroeger zijn nest had gebouwd) en boorde in het zand, waarna hij vervolgens op het zand ging liggen en deed alsof hij doorkroop. Dat wil zeggen: hij sloeg op dezelfde rustige wijze met zijn staart als een stekelbaars die doorkruipt. Hij hield dat een tiental seconden vol en baltste daarna weer verder tegen het vrouwtje. Het lijkt erop (twee studenten hebben ieder ook eens zo'n stekelbaars gehad) dat de stekelbaars de doorkruipbeweging zelfs zonder nest kan uitvoeren. Maar uitvoeren moet hij de beweging wel. Het lijkt niet waarschijnlijk dat hij daarbij een prikkel van buiten registreert die de T-factor doet stijgen, nee, er gebeurt blijkbaar iets in het visje.
De eerstvolgende stap die wij nu gaan doen is kijken of datgene wat er in het dier gebeurt misschien temperatuurgevoelig is. Tussen twee doorkruipen in plaatsen we de stekelbaars dan enige tijd in koud water. Is het proces dat zich in het dier afspeelt beïnvloedbaar door toevoer van koude dan moet dat aan eerder komen, of wat waarschijnlijker is, aan uitstel van de daaropvolgende doorkruip te merken zijn en dan weten we ook weer iets meer over de richting waarin we moeten zoeken.
| |
Waartoe deze regelmaat?
Tot nu toe hebben we ons steeds bezig gehouden met de veroorzaking van de doorkruipritmiek in het gedrag van de
| |
| |
driedoornstekelbaars. In de ethologie is het echter gebruikelijk om je, bij elk gedrag ook altijd af te vragen wat de functie ervan is. Soms is zo'n vraag gemakkelijk te beantwoorden. Neem het waaieren in de broedzorgfase, het is duidelijk waarvoor dat dient. Als het niet gebeurt, gaan eieren dood.
Maar minder duidelijk is het waarom de stekelbaars zo nodig zo precies op tijd door zijn nest moet gaan. Je kunt je in de eerste plaats al afvragen: waartoe moet hij eigenlijk doorkruipen? Doet hij het om de nesttunnel open te houden zodat een kuitrijp wijfje gemakkelijker naar binnen kan kruipen? Maar als je met behulp van de draaitafel een stekelbaars een nest geeft waarin lange tijd niet is doorgekropen zul je zien dat een kuitrijp vrouwtje ook heel gemakkelijk kan inkruipen. En zelfs al zou doorkruipen dienen om de tunnel open te houden dan valt nog niet te begrijpen waarom het zo regelmatig moet geschieden. Het zou bij wijze van spreken voldoende zijn als de stekelbaars doorkroop vlak voordat een wijfje inkruipt.
Je kunt natuurlijk ook zeggen: alle gedrag is ritmisch georganiseerd, ook ons eigen gedrag (denk aan slaap-waak ritme, aan temperatuur-ritmen en dergelijke) en die ritmiek garandeert dat bepaalde, noodzakelijke handelingen op gezette tijden gebeuren. Maar dan nog blijft de vraag bestaan waarom de stekelbaars ritmisch zo strak georganiseerd is. Waartoe kan dat dienen? Of is het slechts een toevallige uitkomst van dat zenuwstelsel? Vragen waarop helaas nauwelijks een antwoord te geven valt.
Het enige wat enig houvast geeft is een vergelijking van het doorkruipritme met ons slaap-waak ritme. Net als bij de stekelbaars resulteert korter slapen dan anders (vergelijk korter in het nest liggen) in eerder weer slaperig worden. Te lang slapen heeft nauwelijks effect, je kunt niet door ‘vooruit’ te slapen bewerkstelligen dat je een nacht later helemaal niet naar bed hoeft. Om te slapen heb je een bed nodig en als het bed vreemd is moet je er eerst aan wennen voordat je erin kunt slapen net zoals een stekelbaars moet wennen aan een nest met een glasplaatje erin. Als het niet anders kan, ben je zelfs nog in staat om helemaal zonder bed te slapen al heb je wel iets nodig om in te zitten of op te liggen (hoewel staande slapen ook mogelijk schijnt te zijn), net zoals de stekelbaars ook zonder nest kan doorkruipen (dat komt tenminste een enkele keer voor). Een heel belangrijke en veelzeggende overeenkomst is wel dat in bed liggen nog niet garandeert dat je slaap binnen haalt zoals bij een stekelbaars
| |
| |
in het nest liggen evenmin garandeert dat zijn T stijgt. En net zoals je, wanneer je slaapt, wakker kunt schrikken en wel blijven liggen (maar dat telt dan niet meer als slaap) kan de stekelbaars tijdens doorkruipen zodanig schrikken van een verbrede glazen uiteinde op zijn nest, dat hij, ook al blijft hij in het nest liggen, niet echt meer doorkruipt. Alle uitwendige omstandigheden kunnen aanwezig zijn om slaap of stijging T-factor te garanderen maar er moet ook iets in de mens, in het dier gebeuren, wil er sprake zijn van slaap of stijging T-factor.
Slaap is zeer noodzakelijk voor een mens, maar er lijkt geen duidelijke reden aanwezig te zijn voor de strakke ritmiek. Je gezondheid lijdt er stellig onder als je onregelmatig slaapt, maar dat is een gevolg van die onregelmatigheid; dat gevolg houdt nog niet in dat je daaruit zou mogen concluderen dat er een duidelijk functionele reden is waarom je zo precies eenmaal in de vierentwintig uur gedurende steeds dezelfde acht uur moet slapen. Misschien moeten we het doorkruipen bij de stekelbaars ook wel als zoiets zien, maar toch blijft deze hoogst precies en blijkbaar zo fijn afgestelde ritmiek een raadselachtig fenomeen.
|
|
