|
| |
| |
| |
7 Neurofysiologische condities
Dit hoofdstuk geeft een kort overzicht van de belangrijkste neurofysiologische mechanismen die een rol spelen in het emotionele proces. Deze mechanismen zullen worden behandeld als de condities voor emotie. In hoofdstuk 3 is aandacht besteed aan de neurale en fysiologische aspecten van de emotionele respons. Wij zullen het in dit hoofdstuk hebben over de zenuwstructuren en de hormonale voorzieningen in de hersenen, waarvan het goed functioneren de conditie vormt voor de normale emotionele respons.
Het overzicht is beknopt en oppervlakkig, deels omdat de literatuur erover zeer uitgebreid, complex en vaak verwarrend is, deels omdat het is toegespitst op een aantal specifieke doeleinden: de afronding van de bespreking over de antecedenten van de emotie; de benadrukking van de biologische grondslag van emotie - hoe belangrijk de rol van de cognitie ook moge zijn; en ten slotte het verkrijgen van inzicht in wat de psychologische theorie over emotie kan opsteken van dit soort neurofysiologische analyses.
Een uitgebreid overzicht van de desbetreffende literatuur is te vinden bij Ruch (1979), Isaacson (1974) en Gray (1982), aan welke auteurs veel in dit hoofdstuk is ontleend.
| |
7.1 Hersenstructuren en emotie
De belangrijkste hersenstructuren die een rol spelen bij de emotie zijn weergegeven in figuur 7.1. De neurofysiologische besturing van het emotionele gedrag en de emotionele beleving, gemotiveerd gedrag inbegrepen, wordt doorgaans beschreven in termen van drie structuren: de hersenstam, waaronder ook bepaalde delen van het diëncefalon, het limbische systeem, en de neocortex. Deze indeling komt in grote lijnen overeen met MacLeans (1970) beschrijving van de ‘hersendrieëenheid’ (‘triune brain’): de protoreptilische hersenen, de protomammale of ‘viscerale’ hersenen, en de neomammale hersenen.
De protoreptilische hersenen staan volgens MacLean in dienst van de elementaire behoeften zoals voedselvoorziening, beschutting, verdediging en voortplanting; ze bevatten de programma's voor soortspecifiek gedrag. Door elektrische stimulatie van de lagere hersenstam is het mogelijk ongeleerde gedragingen op te wekken, of althans fragmenten daarvan. De middenhersenen bevatten onder andere mechanismen die de waaktoestand reguleren en invloed uitoefenen op de prikkeling van hogere hersenstructu- | |
| |
ren. In het diëncefalon wordt de hypothalamus beschouwd als het ‘hoofdganglion’ van de autonome respons, terwijl deze structuur tevens zorg draagt voor de activatie en integratie van soortspecifiek gedrag zoals afweer, voeding en geslachtsverkeer. De hypothalamus is het belangrijkste regelcentrum van de emotionele arousal en van behoeftegebonden gedrag. Hoe belangrijk deze structuur is blijkt uit het feit dat na verwijdering van de volledige hersenschors bij honden of katten het dier nog wel schijnwoede (‘sham rage’) kan vertonen, waarbij minimale stimuli heftige reacties uitlokken zoals blazen, vechten, krabben, en alle autonome verschijnselen die daarmee gepaard gaan (Goltz 1892). Wanneer de hersenstam achter de hypothalamus wordt doorgesneden verdwijnt deze respons, waaruit valt af te leiden dat de hypothalamus het primaire centrum voor de integratie van de affectieve responsen moet zijn (Bard 1928). Voorts bevinden zich in de hersenstam, de middenhersenen en het diëncefalon de oorsprong en een belangrijk deel van het verloop van een aantal belangrijke zenuwbanen, de monoaminebanen, die noradrenaline, dopamine of serotonine afscheiden. Deze lopen door tot in het limbische systeem en tot ver in de neocortex en zijn van
belang voor de regeling van de aandacht, voor de responsbekrachtiging, en voor de stemming (Ungerstedt 1971; Clavier & Routtenberg 1980; McGeer & McGeer 1980).
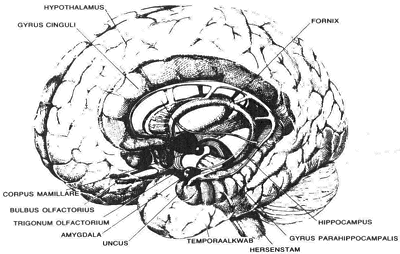
Figuur 7.1 De voornaamste bij emotie betrokken hersengebieden. In aangepaste vorm overgenomen uit F. Snyder, Opiate receptors in de brain, Scientific American, maart 1977.
| |
| |
Ook de basale ganglia - de nucleus caudatus, het putamen en de globus pallidus - worden door MacLean tot de protoreptilische hersenen gerekend. De basale ganglia zijn betrokken bij de organisatie en de regulatie van behoeftebepaalde en doelgerichte actie.
Het limbische systeem is de benaming voor een aantal onderling verbonden structuren, die voor het merendeel behoren tot de fylogenetisch oudere hersendelen. De hoofdstructuren van het limbische systeem zijn de amygdala, de hippocampus, en het septale gebied. Sommige auteurs rekenen er ook de gyrus cinguli en het voorste deel van de thalamus toe (Isaacson 1974). Dit samenstel van structuren wordt verondersteld de responsen te reguleren die door de hypothalamus en de basale ganglia worden georganiseerd, en deze te voorzien van de nodige informatie. Het integreert cognitieve aspecten met actieopdrachten.
De bijdrage van de neomammale hersenen, de neocortex, aan het emotionele proces bestaat uit de cognitieve analyse van relevante informatie. Deze structuur is verantwoordelijk voor het vermogen tot anticipatie, tot subtiele onderscheiding, en voor de analyse van talige informatie. Bovendien draagt ze bij aan de ontwikkeling van plannen en spontane interessen, die van vitaal belang zijn voor het ontstaan van zowel de emoties in engere zin als de emoties van belangstelling of nieuwsgierigheid en het nastreven en realiseren van toekomstplannen.
De bespreking van de neurofysiologische condities zal verder worden uitgewerkt aan de hand van een aantal psychologisch relevante onderwerpen.
| |
7.2 Opwekbaar gedrag
Door elektrische stimulatie van specifieke lokaties in de hersenen kunnen verschillende soorten emotionele en motivationele gedragingen worden opgewekt. Kennelijk vindt de organisatie van het betreffende gedrag op of lager dan de geprikkelde lokaties plaats. De stimulatie blijkt echter alleen het gedrag uit te lokken wanneer er tevens geschikte stimulusobjecten aanwezig zijn; zijn deze er niet, dan blijft het gedrag achterwege. Het is ook mogelijk om door middel van externe stimulatie emotioneel of motivationeel gedrag op te wekken bij dieren waarbij bepaalde hersencentra zijn verwijderd of bepaalde verbindingen in de hersenen zijn doorgesneden. De organisatie van het getoonde gedrag zetelt dan blijkbaar in de nog aanwezige of nog verbonden structuren.
Wanneer bij intacte dieren het laagste gedeelte van de hersenstam wordt gestimuleerd vertonen zij fragmenten van emotioneel gedrag: krabben, bepaalde gelaatsexpressies, geluiden (zie bijv. Delgado & Mir 1969, voor onderzoek bij apen). Bij mensen kunnen ponslaesies leiden tot dwangmatig
| |
| |
lachen of huilen, zonder dat er van emotie sprake is (zie Loeb & Meyer 1969). Dit lijkt erop te wijzen dat de lagere hersendelen elementaire gedragsprogramma's bevatten met een zeer beperkte mate van organisatie. Iets dergelijks blijkt ook uit de responsen op pijnlijke externe stimuli bij dieren waarbij de hersenstam net onder de hypothalamus is doorgesneden. De dieren vertonen afweerreacties - krabben, blazen - die echter slecht gecoördineerd zijn; er is bovendien geen sprake van algemene opwinding. De reactie houdt op zodra de stimulatie wordt beëindigd (Woodworth & Sherrington 1904). Deze reacties worden ‘pseudo-affectieve’ reacties genoemd vanwege de afwezigheid van een algemene opwindingstoestand.
Wanneer daarentegen lokaties in de hypothalamus en het limbische systeem worden gestimuleerd, treden er volwaardige emotionele responspatronen op, met zowel vegetatieve als somatische componenten. Bij Hess (1957) vinden wij de volgende beschrijving van ‘affectieve afweer’ bij een kat, opgewekt door stimulatie van het mediale deel van de hypothalamus. Enkele seconden nadat de stimulatie is begonnen vertoont het dier tekenen van aandacht en opwinding. Het kijkt om zich heen, de haren gaan overeind staan, de pupillen verwijden zich, de kat begint zwaar te ademen. Hij vestigt al zijn aandacht op een persoon in zijn onmiddellijke nabijheid, haalt uit wanneer men hem tracht te benaderen, en zet zich schrap om van het testplatform te springen. Het gedrag van de kat, zegt Hess, kan nauwelijks anders worden opgevat dan als uiting van een gevoel van ernstige bedreiging. Het is tevens te beschouwen als de activatie van een ‘actietendens’: de drempel voor bedreigingssignalen en voor het daaruit resulterende gedrag is verlaagd, en na afloop van de stimulatie blijft het dier enige tijd prikkelbaar en blaast het wanneer er iemand te dicht in de buurt komt. Gedurende de stimulatie vertoont de ontwikkeling van het gedrag een duidelijke samenhang met de situatie: het richt zich tegen mensen en zoekt naar vluchtwegen. Deze adaptieve flexibiliteit is een kenmerkende eigenschap van gedrag dat wordt opgewekt door stimulatie van de hypothalamus, van het limbische systeem, en van sommige delen van de middenhersenen. Kippen die aldus tot de aanval worden geprikkeld pikken duidelijk naar het gezicht van de onderzoeker (Von Holst 1957). Bij apen wekt stimulatie van de centrale grijze stof van de middenhersenen agressie op wanneer het dier omringd is door apen die
een lagere positie bekleden in de sociale hiërarchie, en onderdanigheid wanneer het wordt geconfronteerd met apen van hogere rang (Delgado 1967). Katten richten hun defensieve aanvallen vaker op levende dan op opgezette ratten, vooropgesteld dat beide beschikbaar zijn (Wasman & Flynn 1962).
Door stimulatie van min of meer duidelijk van elkaar te onderscheiden lokaties in de hypothalamus en de middenhersenen kan een groot aantal verschillende emotionele en motivationele gedragingen worden opgewekt. Dit lijkt erop te wijzen dat de diverse gedragssystemen neuro-anatomisch
| |
| |
gezien min of meer afzonderlijke eenheden zijn; misschien geldt dat zelfs voor verschillende componenten van een en hetzelfde gedragssysteem. Sommige onderzoekers zijn echter de mening toegedaan dat de stimulatie die tot affectieve afweer leidt dat alleen maar doet omdat de stimulatie voor het dier onaangenaam is. Dat zulke stimulatie onaangenaam is, is inderdaad vrijwel zeker (zie par. 7.6), maar dat neemt niet weg dat, met gebruik van kleine elektroden, stimulatie van verschillende punten blijkt te leiden tot verschillende soorten gedrag, bijvoorbeeld defensieve afweer of vluchtgedrag, of zelfs tot verschillende patronen van vlucht- of afweergedrag (Romaniuk 1967; Brown, Hunsperger & Rosvold 1969; Wasman & Flynn 1962). Aanvallend prooigedrag (Flynn 1973; Panksepp 1971), agressie tussen mannetjesdieren (Koolhaas 1978; Adams 1971; Kruk e.a. 1979), en defensieve agressie (Brown e.a. 1969) worden opgewekt vanuit duidelijk onderscheidbare laterale en mediale lokaties in de hypothalamus (zie Moyer 1971). De responssystemen vertonen nog een fysiologisch verschil: agressie tussen mannetjesdieren wordt beïnvloed door veranderingen in het testosteronniveau. Dit is niet het geval voor de andere systemen (Bermond 1978).
Door stimulatie van andere laterale zones in de hypothalamus kan appetitief gedrag worden opgewekt: eten, drinken, op hout knagen (bij knaagdieren). Bij mannetjesratten werd een seksuele reactie op daartoe geëigende objecten opgewekt door stimulatie van het achterste gedeelte van de hypothalamus (Caggiula & Hoebel 1966) en van de area preoptica (Van Dis & Larsson 1971).
Zoals gezegd leidt dit soort stimulatie alleen tot de betreffende gedragingen wanneer er een geschikt object aanwezig is. Agressief prooigedrag manifesteert zich alleen wanneer er een potentiële prooi aanwezig is (Flynn 1973), agressie tussen mannetjesdieren alleen in de nabijheid van soortgenoten die zich onderdanig gedragen (Koolhaas 1978). Het effect van hypothalamusstimulatie is niet zozeer dat er een bepaalde reactie wordt opgewekt, maar dat de drempel voor een reactie op een bepaald signaal wordt verlaagd, of dat de aandacht voor bepaalde signalen wordt gewekt. Een kat die in eerste instantie een rat negeert begint belangstelling te tonen zodra ‘de prooiaanvalslokatie’ wordt gestimuleerd. De drempelverlaging is vooral in dit geval duidelijk. Wanneer men de stimulatie sterker laat worden, vergroten zich de sensorische oppervlakken in gezicht en voorpoten van waaruit tactiele prikkels de relevante componenten van het prooigedrag opwekken, respectievelijk het openen van de bek en het uitslaan van de klauwen (Flynn 1973). Operatieve beschadiging van de hypothalamus veroorzaakt juist een toestand van prikkelveronachtzaming (‘sensory neglect’): het dier vertoont geen oriëntatiereactie op aangeboden prikkels. Dit duurt tot enige tijd na de operatie (Marshall & Teitelbaum 1974). Al deze gegevens wijzen erop dat door stimulatie een volledig gedragssysteem
| |
| |
wordt geactiveerd - een actietendens wordt opgewekt - en niet een op zichzelf staande respons. Dat dit het geval is blijkt eveneens uit het feit dat door stimulatie van ‘prooiaanvalslokaties’ net onder het drempelniveau de reactie op potentiële prooien gefaciliteerd blijft tot enige tijd na de beëindiging van de stimulatie, en uit het feit dat het geprikkelde dier lijkt te zoeken naar een geschikte prooi of tegenstander wanneer deze er niet is (Flynn 1973).
Over het algemeen worden door hersenstimulatie dus gedragssystemen en niet afzonderlijke gedragingen opgewekt. Bij stimulatie van de affectieve afweerlokaties produceren zwakke prikkels aandacht en exploratief gedrag. Bij sterkere stimulatie gebeurt hetzelfde wanneer er geen potentieel dreigende stimuli aanwezig zijn; is dit wel het geval dan is de respons een aanvals- of vluchtreactie. Voor appetitief gedrag - eten, drinken, knagen, seksuele activiteit - geldt dezelfde algemene regel: het geëigende gedrag wordt vertoond wanneer er geschikte objecten aanwezig zijn; zijn deze er niet dan is de respons aandacht, exploratie (bij ratten, vooral snuffelen), en rondlopen (Panksepp 1982). Welk type gedrag wordt opgewekt door stimulatie van een bepaalde lokatie is niet louter van de lokatie afhankelijk. Dezelfde stimulatie die aanzet tot eten kan maken dat het dier zijn jongen gaat versjouwen of op hout gaat knagen, wanneer men het voer vervangt door een van deze laatste objecten (Valenstein, Cox & Kakolewski 1970). Bovendien hangt het soort gedrag dat wordt opgewekt niet alleen af van de externe stimulatie en de plaatsing van de elektroden, maar evenzeer van de aard en de achtergrond van het individu (Panksepp 1982; Ploog 1970). Alleen bij ratten die van nature muizendoders zijn zal elektrische stimulatie het dier aanzetten tot het doden van een muis (Panksepp 1971), net zoals het alleen mogelijk is om door middel van hersenstimulatie een driftaanval op te wekken bij mensen die ook al eerder dergelijke driftaanvallen vertoonden. Deze observaties brachten Valenstein (1969) tot de hypothese dat door stimulatie van belonende zones in de hypothalamus die motivationele systemen worden geactiveerd die op het moment van de stimulatie ‘over-heersend’ zijn.
Of, zoals Panksepp (1982) het formuleert: stimulatie activeert verlangen, of verwachting, of bevrediging, of om het even welk doel dat op een bepaald moment bij het individu opkomt onder invloed van prikkels van buitenaf en onder invloed van zijn eigen innerlijke gesteldheid.
De hypothese van de overheersende respons, of van de ‘verwachting’, kent echter zijn beperkingen. Niet alle soorten gedrag kunnen met evenveel gemak in alle lokaties worden opgewekt. Prooigedrag is bijvoorbeeld slechts op te wekken in zeer specifieke lokaties. Dat neemt niet weg dat de stelling dat hersenstimulatie actietendensen of gedragssystemen in werking zet, met de daarbij behorende toestanden van gedragsbereidheid en -flexibiliteit, lijkt te gelden voor alle structuren boven de hersenstam.
Zoals gezegd kan er ook gedrag worden opgewekt vanuit diverse limbi- | |
| |
sche en andere lokaties. Dit soort gedrag vertoont over het algemeen de eerder genoemde eigenschappen; bij mensen leidt stimulatie van deze structuren dikwijls tot welomschreven emotionele ervaringen. Door stimulatie van de amygdala kunnen bijvoorbeeld oriëntatiereacties en angstige en boze responsen worden opgewekt, met zowel gedragsmatige als autonome componenten (Ursin & Kaada 1960; Fonberg 1979). Door stimulatie opgewekt aanvalsgedrag komt geleidelijk op gang, langzamer dan aanvalsgedrag voortkomend uit stimulatie van de hypothalamus, en verdwijnt nog langzamer na het beëindigen van de prikkeling (King, naar Isaacson 1974). In het onderzoek bij mensen zijn woedeaanvallen geconstateerd bij patiënten met tumors in de temporaalkwab, temporale epileptische haarden, en temporale laesies. De aanvallen worden veelal uitgelokt door onbeduidende voorvallen en gaan dikwijls vergezeld van aura-belevingen (Poeck 1969). Bij patiënten met temporale laesies leidt stimulatie van de amygdala tot een als oncontroleerbaar gevoelde opwelling van woede en agressie, vergelijkbaar met wat de patiënten zeggen te voelen tijdens een spontane driftaanval (Mark & Ervin 1970; King 1961).
Stimulatie van andere zones (caput van de nucleus caudatus, Heath 1976; Hassler & Riechert 1961; voorste deel van de thalamus, zie Poeck 1969) kan een reactie teweegbrengen waarbij de patiënt onbedwingbaar moet lachen of glimlachen, en vaak hevige gevoelens van plezier en geamuseerdheid heeft: ‘De patiënt lijkt alles grappig te vinden’ (Hassler & Riechert 1961). Kunstmatig opgewekt gedrag heeft echter niet in alle gevallen een eigen en toepasselijke emotionele inhoud. We hebben vermeld dat irriterende ponslaesies soms niet-emotioneel lachen en huilen veroorzaken. Hoger gelegen laesies van de zenuwbanen tussen thalamus en pons (bij pseudobulbairparese) brengen oncontroleerbare lach- of huilaanvallen teweeg bij ongeveer de helft van de patiënten. Deze aanvallen worden vaak uitgelokt door onbetekenende stimuli, of helemaal nergens door, en de patiënt heeft er doorgaans geen enkel gevoel van verdriet, vreugde of plezier bij. Soms is er zelfs gelijktijdig sprake van volkomen ongelijksoortige emoties (Bechterev 1894; zie Poeck 1969; Rinn 1984). Bij sommige patiënten komen zowel lach- als huilaanvallen voor, bij andere slechts een van de twee (Poeck 1969).
Stoornissen in bepaalde zones (neurotransmitterstoornissen in de basale ganglia bij de ziekte van Parkinson) kunnen daarentegen tot gevolg hebben dat bepaald emotioneel gedrag, gelaatsexpressies in het bijzonder, gestoord wordt. Bij patiënten met deze stoornissen is sprake van ‘mimetische aangezichtsverlamming’, dat wil zeggen zij missen het vermogen tot spontane gelaatsexpressie maar zijn wel in staat hun aangezichtsspieren opzettelijk te bewegen (zie Rinn 1984).
Uit de verschijnselen van door elektrische stimulatie opgewekt gedrag, en uit de effecten van hersenlaesies, kunnen verscheidene punten worden
| |
| |
afgeleid die van belang zijn voor het onderzoek naar emotie.
In de eerste plaats zijn er veel elementaire gedragingen, met inbegrip van de gelaatsexpressies, die kennelijk op het subcorticale niveau zijn vastgelegd; sommige zelfs onderin de hersenstam. Dat wil zeggen dat het ongeleerde gedragswijzen zijn. Bovendien is de regulatie van dit gedrag verschillend van die van willekeurig gedrag. Dat laatste blijkt vooral uit de zojuist genoemde mimetische aangezichtsverlamming en uit de tegenhanger daarvan, de ‘willekeurige aangezichtsverlamming’, waarbij de spontane gelaatsexpressie behouden blijft en verlamming optreedt van de willekeurige beweging. Willekeurige aangezichtsverlamming komt voor bij laesies van het corticale gedeelte van de motore baan van cortex naar hersenstam (zie bijv. Monrad-Krohn 1924; Rinn 1984).
In de tweede plaats blijkt uit de besproken verschijnselen dat er verschillende niveaus van organisatie van het betrokken emotionele en motivationele gedrag bestaan. Op de meer centrale niveaus zijn de gedragswijzen georganiseerd tot systemen die worden gekenmerkt door adaptieve flexibiliteit en door specifieke voorzieningen voor zintuiglijke ontvankelijkheid; subjectief gaat hun activatie gepaard met gevoelens, en met de ervaring dat men door drang tot het gedrag wordt aangezet.
In de derde plaats wijzen bepaalde resultaten erop dat de verschillende gedragssystemen neuro-anatomisch gescheiden zijn. Sommige ervan staan duidelijk los van elkaar terwijl er gedragsmatig een zeker verband bestaat, zoals defensieve agressie, agressief prooi gedrag en agressie tussen mannetjesdieren (Moyer 1971). Andere zijn nauwer met elkaar verbonden dan in de emotietheorie tot nu toe werd aangenomen, bijvoorbeeld ontsnappingsgedrag en defensief aanvalsgedrag (varianten van vrees en woede), die samen het ‘defensieve afweersysteem’ vormen (Hess 1957; Gray 1982), en de gedragsaspecten die in Panksepps visie onder de noemer ‘verlangen’ of ‘verwachting’ vallen (Panksepp 1982).
| |
7.3 Activeringsmechanismen
Een van de steeds terugkerende thema's in de psychologie van de emotie is de relatie tussen emotionele verschijnselen en de intensiteit van gedrag. In de voorgaande hoofdstukken is aandacht besteed aan de begrippen arousal en activatie, en aan de vraag of de verschijnselen een eendimensioneel concept rechtvaardigen. Vanuit descriptief oogpunt leek het juister verschillende categorieën te onderscheiden (par. 3.9). Ook de neurofysiologische bevindingen vragen om een indeling in een aantal verschillende activeringssystemen die gerelateerd zijn aan verschillende gedragsverschijnselen of fysiologische verschijnselen.
Het systeem dat traditioneel verantwoordelijk wordt geacht voor activa- | |
| |
tieverschijnselen is de reticulaire formatie van de middenhersenen. Elektrische stimulatie van deze structuur leidt bij een slapend dier tot ontwaken, gedragsmatige en autonome oriëntatiereacties, en desynchronisatie in het eeg (Magoun 1958; Lindsley 1951). Wanneer de reticulaire formatie wordt vernietigd raakt het dier in coma, of wordt het eeg-patroon gesynchroniseerd, of beide. Onder normale omstandigheden wordt de reticulaire formatie geactiveerd door stimulusveranderingen of door informatie-invoer in het algemeen, dat wil zeggen nieuwe stimuli, gedragsonderbreking, of stimuli die in strijd zijn met de verwachtingen. Zelf bevordert de reticulaire formatie de verwerking van zintuiglijke informatie in de hersenschors en activeert zij de autonome arousalrespons en de gedragsmatige waakzaamheid door middel van mechanismen in het voorste gedeelte van de hypothalamus. Volgens Routtenberg (1968) is de functie van het systeem waar de reticulaire formatie deel van uitmaakt de regulering van de algemene responsbereidheid of ‘drijfveer’ (Hulls ‘drive’). Pribram en McGuinnes (1975) gebruiken voor dit systeem de term ‘arousal’. De functie die zij eraan toeschrijven is de interruptie van gedrag dat aan de gang is, de registratie van gebeurtenissen in het bewustzijn, en het inleiden van cognitieve assimilatie - de vorming van ‘neuronale modellen’.
Een onbeschadigde reticulaire formatie blijkt echter niet de enige voorwaarde te zijn voor verschijnselen als ontwaken en eeg-desynchronisatie, of voor gedragsmatige responsen in het algemeen. Dieren met reticulaire laesies raken niet altijd in coma; soms vertonen ze nog steeds een laag gevolteerde, snelle activiteit op het eeg, en ze zijn soms ook nog in staat tot het verrichten van ingewikkelde taken. Het kan ook voorkomen dat dieren hun reactievermogen kwijtraken door andere laesies, terwijl de reticulaire formatie intact is. Door beschadigingen in het achterste gedeelte van de hypothalamus kunnen dieren lethargisch worden; stimulatie leidt dan wel tot een reactie, maar de dieren vallen daarna meteen weer in slaap (zie Routtenberg 1968 voor literatuurverwijzingen). Beschadiging van de grote catecholaminebanen in de middenhersenen - het dopaminerge nigrostriaire systeem en het dorsale noradrenalinesysteem - veroorzaken eveneens lethargie en overmatig slapen (Jouvet 1972).
Deze gegevens lijken erop te wijzen dat er sprake is van een tweede activeringssysteem, door Routtenberg ‘arousal twee’ genoemd, door Pribram en McGuinnes aangeduid met de term ‘activatie’. Volgens deze laatsten is de functie hiervan de preparatie van responsmechanismen, in tweeerlei opzicht: het vasthouden van de verwachtingen, en het tot stand brengen van somatomotorische paraatheid. De functie van het systeem is dus de activatie van intentionele gerichtheid, van tonische, waakzame paraatheid, en tevens van specifieke intentionele handelingen. Dit systeem omvat de eerder genoemde dopaminerge nigrostriaire baan en de basale ganglia, waarvan bekend is dat zij een rol spelen in de organisatie van willekeurige
| |
| |
beweging, in de handhaving van de actieve lichaamshouding, en in de organisatie van met behoeftebevrediging verbonden gedrag zoals eten en drinken. Het systeem omvat alle of een deel van de laterale hypothalamuslokaties vanwaar appetitief gedrag kan worden opgewekt (zie par. 7.2). Zoals gezegd bestaan er aanwijzingen dat bij dit soort gedragsopwekking sprake is van activatie van responssystemen die overheersend zijn vanwege de aanwezige stimuli of de voorgeschiedenis van het subject. Pribram en McGuinness noemen dit systeem ook wel het startsysteem (‘go’ system).
De dopaminerge nigrostriaire baan, de naam zegt het al, loopt naar het corpus striatum en naar de basale ganglia. Beschadiging ervan veroorzaakt lethargie. De normale functie van deze structuur lijkt de niet-specifieke activatie van de basale ganglia te zijn, resulterend in het zojuist beschreven tonische niveau van paraatheid. Specifieke stimulusaanvoer via andere routes zorgt voor de voorbereiding van specifieke handelingen, met die tonische achtergrond als basis. Waarschijnlijk kan de dopaminerge baan dit soort aspecifieke activatie produceren omdat dopamine in de basale ganglia niet zozeer als neurotransmitter fungeert, maar als neuromodulator. Met andere woorden, het moduleert de werking van de neurale systemen die door de echte neurotransmitters (in dit geval acetylcholine) gevoed worden (Fuxe, geciteerd in Pribram 1977).
Het door Pribram en McGuinness gemaakte onderscheid tussen arousal en activatie is in dit verband van aanzienlijk belang, niet alleen omdat het opheldering heeft gegeven omtrent deze begrippen, maar ook omdat eruit is af te leiden dat er twee verschillende systemen ten grondslag liggen aan het optreden van emoties in engere zin enerzijds, en van motivationeel gedrag of verlangens anderzijds. In het eerste geval is er sprake van ‘arousal’, in de betekenis die Pribram en McGuinness eraan geven, dat wil zeggen de activatie van een gedragsonderbrekend en oriënterend (‘stop-and-see’) systeem, in het tweede geval is er sprake van ‘activatie’, waartoe actietendensen zoals wij die eerder beschreven hebben, tendensen om te vechten of te vluchten bijvoorbeeld, behoren.
Pribram en McGuinness (1975) onderscheiden een derde systeem in de regulatie van activiteit, het ‘effort’-systeem met de hippocampus als centrale component. Dit systeem is verantwoordelijk voor willekeurige concentratie en voor de organisatie van reactiewijzen die afwijken van het op grond van de aanwezige stimuli gebruikelijke of meest natuurlijke reactiepatroon. Hiertoe behoort onder andere de gedragsinhibitie, en we zullen straks in dat verband het systeem nader bespreken. We komen eveneens later (par. 7.6) terug op de hormonen die als neuromodulatoren optreden, acth en de corticosteroïden; deze stoffen verhevigen de alertheid, waarschijnlijk doordat ze de werking van de hippocampus stimuleren.
De activeringssystemen zijn in complexe relaties verwikkeld met andere systemen waarvan ze informatie ontvangen en die zij op hun beurt beïn- | |
| |
vloeden met hun output. Wij kunnen dit illustreren aan de hand van de invloed van en naar het achterste gedeelte van de hypothalamus, dat naar alle waarschijnlijkeid deel uitmaakt van zowel het arousalsysteem als het effortsysteem. Zoals gezegd veroorzaken laesies in het achterste gedeelte van de hypothalamus slaperigheid en lethargie. Stimulatie van deze structuur leidt niet alleen tot sympathische arousal en eeg-desynchronisatie, maar ook tot spierbewegingen. Dit laatste effect lijkt onafhankelijk te zijn van het eerste (Gellhorn 1968). Het lijkt erop dat dit de manier is waarop de actieve emoties geactiveerd gedrag veroorzaken. De activiteit van het achterste gedeelte van de hypothalamus is zelf afhankelijk van feedback van de spieren en de gewrichten. Door curare wordt die activiteit verminderd en daardoor ook, indirect, de corticale arousal. Spieractiviteit blijkt dus de arousal te verhogen en zichzelf in stand te houden (Gellhorn 1968). Dit zou kunnen verklaren waar het opwekkende effect van lichaamsbeweging vandaan komt.
Er zijn nog andere structuren betrokken bij de activatie van gedrag en bij het optreden van emotionele responsen. De belangrijkste hiervan zijn de amygdala en de frontaalkwabben. Volgens Pribram en McGuinness maken deze structuren deel uit van het arousalsysteem. Zij beschouwen de amygdala als de zetel van de voornaamste regulatiemechanismen van dat systeem. Wij zullen ze echter onder een iets andere noemer bespreken.
| |
7.4 Evaluatiefuncties
Bilaterale vernietiging van de temporaalkwabben veroorzaakt bij apen drastische veranderingen in de emotionele responsiviteit. De apen worden rustig en mak, en tonen zich onverschillig tegenover de normale vreeswekkende stimuli zoals slangen of vreemden. Bovendien verliezen ze hun normale agressiviteit. Tegelijkertijd kunnen ze hyperseksueel gedrag gaan vertonen - dat is althans het geval bij dieren in gevangenschap. Dit betekent dat ze geen onderscheid meer maken tussen geschikte en ongeschikte seksuele objecten, en bijvoorbeeld proberen te paren met een kat of een konijn. Ook vertonen ze vaak uitgesproken oraal gedrag, met de dwangmatige neiging alles wat ze zien in hun mond te steken. Dit ‘Klüver-Bucy-syndroom’ (Klüver & Bucy 1937), of althans duidelijke aspecten ervan, is ook aangetroffen bij mensen met bilaterale beschadiging van de temporaalkwabben. De stemming van dit soort patiënten is over het algemeen onverschillig, soms is er sterke oraliteit, en er kan sprake zijn van dwangmatig, gulzig eten. Ook zijn ongeremde seksuele uitingen geobserveerd, waarbij de patiënt zich niet stoorde aan de aanwezigheid van andere personen (Poeck 1969).
Duidelijke aspecten van het syndroom zijn waar te nemen wanneer de
| |
| |
beschadiging is beperkt tot de amygdala en het omringende weefsel (Schreiner & Kling 1953). Bij honden en katten verdwijnt de agressiviteit; ook bij agressieve dieren zoals de lynx is dat het geval. Verwijdering van de amygdala bij apen en bij mensen leidt, te oordelen naar klinische waarnemingen, tot ernstige aantasting van de gevoeligheid voor sociale signalen. In kolonies die in gevangenschap leven verliezen mannelijke resusapen bij wie de amygdala zijn verwijderd hun status in de sociale hiërarchie. Dit is vooral te wijten aan het feit dat ze hun gedrag niet meer afstemmen op de specifieke situatie (Rosvold e.a. 1954). In het wild worden het sociale outcasts. Ze raken in ongebruikelijke sociale conflicten verwikkeld en trekken zich ten slotte uit de groep terug (Dicks e.a. 1969). Honden zonder amygdala verliezen hun aanhankelijke gedrag jegens hun baas, met het gevolg dat tevens al het gedrag dat wordt bekrachtigd door de goedkeurende aanmoediging van de baas achteruit gaat (Fonberg 1981).
Isaacson (1974) formuleert het als volgt: dieren waarbij de amygdala zijn weggenomen schijnen, althans in een beschermde omgeving, ‘tevreden te zijn met het lot dat hun toevalt’. Ze trekken zich weinig aan van onverwachte gebeurtenissen en reageren daar nauwelijks op (Bagshaw e.a. 1972). Fonberg (1981) beschrijft haar honden met dorsomediale amygdalalaesies als onverschillig en emotieloos. Toch zijn zulke dieren emotioneel niet volledig afgestompt. Er treden bijvoorbeeld wel aandachtsreacties op als de stimuli voldoende belonend zijn (Pribram, Douglas & Pribram 1969), en er wordt een angstige of agressieve reactie opgewekt wanneer het belang van de gebeurtenis niet genegeerd kan worden, bijvoorbeeld wanneer het dier wordt aangevallen door soortgenoten (Rosvold e.a. 1954). Dieren zonder amygdala blijven op dit soort ongebruikelijke gebeurtenissen reageren alsof het nieuwe stimuli zijn, ook wanneer ze er herhaaldelijk mee geconfronteerd worden (Schwartzbaum e.a. 1961).
De functie die zowel bij dieren als bij mensen met temporale laesies het meest lijkt te zijn aangetast is het vermogen om gemotiveerd gedrag aan te passen aan de veranderende omstandigheden. Dit wordt met name geïllustreerd door verschijnselen zoals dwangmatig eten en hyperseksualiteit: de dieren en de patiënten blijven doorgaan wanneer normale subjecten zouden ophouden, ze staan machteloos tegenover aanlokkelijke stimuli. Er zijn ook gevallen bekend van extreem agressief gedrag als reactie op lichte provocaties, vermoedelijk ten gevolge van temporaal letsel (Mark & Ervin 1970).
Al deze gegevens doen vermoeden dat het intacte organisme over evaluerende functies beschikt die de gebeurtenissen beoordelen op hun hedonische of regulerende relevantie, en die activerende of inhiberende informatie leveren aan de hypothalamussystemen die de eigenlijke emotionele respons besturen. Pribram en McGuinness (1975) beschouwen de amygdala dan ook als het belangrijkste regelcentrum van het arousalsysteem dat
| |
| |
gevoelig is voor veranderingen in stimulusomstandigheden. Het is derhalve te beschouwen als het voornaamste regelcentrum van de emoties in engere zin.
In de hierboven beschreven observaties van subjecten met temporale laesies is vooral de nadruk gelegd op het afstompende effect dat door dit soort laesies wordt teweeggebracht, en, als conclusie daaruit, op het activerende effect van de evaluatieve functies. Dat activerende effect wordt echter geneutraliseerd door tegenovergestelde, dat wil zeggen remmende effecten. Dat blijkt uit het feit dat temporale laesies zowel kunnen leiden tot hyperseksualiteit en oraliteit als tot emotionele vervlakking. Men blijkt in de amygdala dan ook verschillende gedeelten te kunnen onderscheiden die elkaar in veel opzichten antagoneren (Kaada 1967; Fonberg 1979). Laesies in bepaalde gebieden maken dat dieren uitermate agressief worden in plaats van kalm en passief (Wood 1958; Fonberg 1979). In sommige gebieden in de amygdala veroorzaakt elektrische stimulatie arousal en agressieve responsen, in andere arousalafname, angst, of seksuele opwinding (Ursin & Kaada 1960). Bij mensen is gebleken dat stimulatie van hét kerngebied van de amygdala angst teweegbrengt (Heath, Monroe & Mickle 1955; Chapman 1958). Op grond van de waargenomen reciproke effecten hebben Pribram en McGuinness (1975) een systeem van reciproke besturing gepostuleerd door middel waarvan een uiterst genuanceerd gereguleerde afstemming van de hypothalamusarousal mogelijk is. Deze afstemming is, moet men aannemen, gebaseerd op elders gegenereerde informatie.
Zoals gezegd maken bepaalde gebieden van de frontaalkwabben volgens Pribram en McGuinness deel uit van het arousalsysteem in de door hen gebruikte betekenis. Verwijdering van de frontaalkwabben veroorzaakt bij apen een aanzienlijke afname van het eigen initiatief, de dieren volharden niet meer in doelgerichte handelingen en hun agressieve responsen op frustratie zijn aanzienlijk verminderd (Jacobsen 1936). Dieren met letsel aan de frontaalkwabben tonen aandacht voor elke verandering in de omgeving, maar zijn niet in staat een instelling op gebeurtenissen in de toekomst te handhaven, zoals nodig is bij taken die om een uitgestelde respons vragen (zie Rosenkilde 1979 voor een overzicht). Bij mensen resulteert letsel aan de frontaalkwabben in gebrek aan interesse, initiatief en doelbewuste regulatie van het gedrag in moeilijke situaties (Powell 1979; Kleist gebruikt de term ‘Antriebsschwäche’). De stemming van de patiënt is onverschillig of vlak eufoor. Net als in het genoemde onderzoek reageert de patiënt op allerlei irrelevante stimuli waaraan, ondanks herhaaldelijke presentatie, geen gewenning optreedt. De regulatie van activiteit door plannen die het subject nog wel kan verwoorden blijkt ernstig te zijn verstoord. Luria (1973) trekt uit deze observaties de conclusie dat ‘de frontaalkwabben bij de mens een directe bijdrage leveren aan de toestand van verhoogde activatie waarmee alle vormen van bewuste activiteit gepaard gaan’. Deze ‘ver- | |
| |
hoogde activatie’ lijkt essentieel te zijn voor het hebben van interessen, alsmede voor de meer subtiele emoties. Luria (1969) beschrijft het frontaal-kwabsyndroom als vernauwing van de interesse, en desintegratie van de hogere vormen van emotionele
beleving. De betrokken functies lijken zelfs van wezensbelang te zijn voor de meer elementaire emoties. Door prefrontale lobotomie is het mogelijk onbehandelbare pijn weg te nemen - de patiënt is zich nog wel bewust van de pijn maar heeft er geen last meer van - alsook hevige angst en dwangverschijnselen (Powell 1979). Een dergelijke behandeling leidt echter tevens tot de eerder genoemde verschijnselen: apathie, interesseverlies en onvermogen tot actieplanning.
Net als de amygdala vertonen de frontaalkwabben een functionele differentiatie. Orbitofrontale laesies kunnen bij apen verhevigde vrees- en woedereacties veroorzaken (Butter & Snyder 1972), en bij mensen verregaande emotionele ongeremdheid, uitbarstingen van razernij, en ingrijpende karakterwijzigingen. Planningsstoornissen en apathie treden vooral op bij laterale laesies (Luria 1969,1973). De frontaalkwabben bevatten, net als de amygdala, antagonistische, reciprook werkende gebieden die respectievelijk orbitofrontaal en frontolateraal zijn gelokaliseerd (Pribram & McGuinness 1975).
| |
7.5 Inhibitie- en regulatiefuncties
Er spelen verschillende soorten inhibitiemechanismen een rol bij de antagonistische, reciproke besturing van een aantal functionele systemen. Een hiervan, de tweeledige besturing van de emotionele arousal door de reciproke circuits van amygdala en frontaalkwabben, is in de vorige paragraaf behandeld. Van een soortgelijke tweeledige besturing is sprake in de wijze waarop de hypothalamus functioneert. Stimulatie van het achter- en middengedeelte van de hypothalamus veroorzaakt voornamelijk sympathische responsen, van het voorste en laterale gedeelte parasympathische responsen. Op grond hiervan maken Hess (1957) en Gellhorn (Gellhorn & Loofburrow 1963) het onderscheid tussen ‘ergotrope’ en ‘trofotrope’ zones in de hypothalamus, die samen een wisselend evenwicht teweegbrengen in de besturing van de autonome functies. Er lijkt tevens sprake te zijn van een zekere reciproke controle in de aanzet tot en de beëindiging van appetitief gedrag zoals eten en drinken; dit speelt zich eveneens gedeeltelijk op het niveau van de hypothalamus af. De aanzet tot eten of drinken wordt gereguleerd door mechanismen in de laterale hypothalamus of de aangrenzende weefsels onder de hypothalamus. De beëindiging ervan wordt bepaald door mechanismen die onder invloed staan van ventromediale hypothalamusprocessen. Misschien staan deze laatste in verband met de afremming van het plezier in eten (zie Grossman 1979 voor een overzicht). Er zijn
| |
| |
aanwijzingen dat deze twee mechanismen een wederzijds inhibitoir effect hebben (zie Pribram 1980).
We hebben tot nu toe gesproken over impulsregulatie door de reciproke werking van inhiberende en activerende mechanismen. Er kan echter ook sprake zijn van directe inhibitie van emotioneel gedrag of emotionele impulsen. We hebben al gezien dat verwijdering van de gehele hersenschors bij honden en katten hevige schijnwoede veroorzaakt als reactie op lichte provocaties (Bard & Mountcastle 1948). Hieruit blijkt dat de cortex een remmende invloed uitoefent op de onderliggende structuren. Deze inhibitoire beheersing speelt ook een rol bij bewuste zelfbeheersing en bij de beïnvloeding van spontane gelaatsexpresies door cultuurbepaalde uitingsregels (Rinn 1984). Wanneer de inhibitoire corticale beheersing wordt verzwakt, bijvoorbeeld door alcoholgebruik, kan het subject ongeremd gedrag gaan vertonen.
Mechanismen die verantwoordelijk zijn voor gedragsinhibitie bevinden zich in de hersenstam en in het limbische systeem, en ook in de hogere corticale structuren. Hess (1957) vond dat prikkeling van de basale ganglia - nucleus caudatus en corpus striatum - de spontane activiteit onderbrak en onbeweeglijkheid veroorzaakte. Delgado (1975, 1979) constateerde dat door dergelijke stimulatie agressief gedrag en eetgedrag werden onderbroken, waarbij het niet uitmaakte of deze gedragingen spontaan waren of waren opgewekt door stimulatie van andere hersenstructuren. Volgens Delgado kunnen door stimulatie van specifieke lokaties bepaalde gedrags-systemen selectief worden geremd. Om een voorbeeld te geven: een dier dat onder het eten geprikkeld wordt door een elektrode op een specifieke plaats, houdt op met kauwen en verliest zijn belangstelling voor het voedsel of keert zich er met een grimas van afschuw vanaf. Het wordt echter niet onbeweeglijk. Een aanvallende stier die wordt geprikkeld op een bepaald, vermoedelijk agressie-inhiberend punt in de hersenen staakt zijn aanvalsgedrag en begint andere bewegingen te maken (Delgado 1964). De interpretatie van deze bevindingen in termen van specifieke inhiberende circuits is echter aangevochten. Stimulatie zou bijvoorbeeld incompatibele gedragspatronen kunnen opwekken (Valenstein 1973).
Laesies in het septale gebied veroorzaken een hyperactieve reactie op elementaire schrikstimuli zoals lichtflitsen, plotseling geluid en elektrische schokken. De schrikreacties zijn aanzienlijk verhevigd (Lubar e.a. 1970) hetgeen erop lijkt te wijzen dat de normale schrikreactie onder inhibitoire controle staat. Hyperactiviteit komt ook voor bij de zogenaamde ‘septale woede’ die ratten en misschien ook andere diersoorten vertonen na septale laesies (Brady & Nauta 1955). Septale hyperactiviteit is echter slechts van tijdelijke aard.
Stimulatie van de hippocampus veroorzaakt stopreacties: activiteit die aan de gang is wordt onderbroken en het subject wordt bewegingloos (Mac- | |
| |
Lean 1957; Kaada, Jansen & Andersen 1953). Hippocampusbeschadigingen leiden daarentegen tot een wisselvallige activiteitstoename. Er wordt met meer handelingen begonnen, op meer nieuwe gebeurtenissen gereageerd, de reacties op onverwachte en onaangename gebeurtenissen zijn sterk verhevigd, maar de respons blijft vluchtig en van korte duur (Isaacson 1974). Dieren waarbij de hippocampus is weggenomen plegen te volharden in geleerde activiteiten, of in eenmaal gekozen tactieken, ook wanneer de omstandigheden veranderen. Het vermogen tot spontane afwisseling en de neiging om telkens nieuwe plekken te exploreren blijken ernstig te zijn aangetast (Kimble 1968).
Uit dit alles valt af te leiden dat bij beschadiging van de hippocampus en het septale gebied een bepaalde functie van gedragsinhibitie wegvalt, en dat zo'n functie onder normale omstandigheden wel werkzaam is. Ook uit andere activiteiten die door hippocampus- of septumlaesies worden aangetast blijkt een dergelijke functie. Tot deze activiteiten behoren passief vermijdingsgedrag, dat wil zeggen nalaten om naar een aantrekkelijke stimulus toe te gaan vanwege de bestraffing die daarop volgt; uitstel van de respons bij het leren met uitgestelde bekrachtiging, dat wil zeggen het dier wordt beloond als het pas na enige tijd op een stimulus reageert; en responsextinctie, dat wil zeggen het dier moet ophouden met reageren omdat het niet meer beloond wordt. Al deze drie reactiewijzen verslechteren (zie Gray 1982 voor een overzicht). In alle drie de gevallen lijkt er sprake te zijn van verzwakking van een ‘gedragsinhibitiesysteem’ dat gevoelig is voor geconditioneerde bestraffing, of voor ‘frustrative non-reward’, het uitblijven van een verwachte beloning; dit is althans de hypothese van Gray (1982). Volgens deze hypothese is de werking van dit systeem hetzelfde als het angstmechanisme: angst is een inhibitoire respons op strafsignalen en op het uitblijven van verwachte beloning.
De inhibitie hoeft echter niet verzwakt te zijn door beschadiging van de inhibitiemechanismen als zodanig. Het kan ook het gevolg zijn van beschadiging van de structuren die de inhibitiemechanismen van informatie voorzien. Dit is een interessante interpretatie die in verschillende theorieën naar voren komt.
De hippocampus lijkt betrokken te zijn bij bepaalde cognitieve functies, zoals het geheugen en de constructie van cognitieve representaties van de omgeving. Door hippocampusletsel veroorzaakte gedragsstoornissen hebben dikwijls duidelijk cognitieve aspecten. Dieren waarbij de hippocampus is verwijderd vertonen bijvoorbeeld niet alleen de eerder genoemde gebreken, ze zijn ook niet in staat om de aangeboden informatie op adequate wijze te gebruiken, of doen daar althans erg lang over. Ze gedragen zich alsof de situaties waarmee ze herhaaldelijk geconfronteerd worden nieuw voor hen blijven (zie O'Keefe & Nadel 1978 en Gray 1982 voor een bespreking). De gedragsstoornissen blijken vooral wanneer er sprake is van onze- | |
| |
kerheid die vereist dat het gedrag steeds wordt aangepast (Isaacson 1974; Pribram & Isaacson 1975). Voorts wijst de elektrische activiteit in de onbeschadigde hippocampus erop dat deze een rol speelt bij het onderkennen van nieuwe stimuli en van overeenstemming of strijdigheid, match of mismatch, met verwachtingen (zie Pribram & Isaacson 1975 en Gray 1982 voor een overzicht). Dit soort informatie heeft onmiskenbaar belang voor de emotionele besturing.
Volgens Gray is een van de voornaamste functies van dit systeem het aanpassen van de gedragsinhibitie en de aandacht aan het onderkennen van nieuwe en van de verwachting afwijkende stimuli. Volgens Pribram (Pribram & McGuinness 1975; Pribram & Isaacson 1975) dient het er vooral toe de onmiddellijke reactie op stimulatie te sturen, om te zien of die reactie van toepassing is of dat er beter een andere, minder dominante reactiemodus kan worden aangewend. Tot die modi behoren inhibitie, niet-specifieke defensieve bereidheid, angst en concentratie, dat wil zeggen de organisatie van willekeurige, doorredeneren bepaalde activiteit. Zoals gezegd noemen zij dit systeem het effortsysteem.
Het gedragsinhibitie- of effortsysteem kan zijn functies alleen uitoefenen wanneer het toegang heeft tot relevante informatie. Deze informatie is waarschijnlijk afkomstig van anatomisch verbonden gebieden, met name de gyrus cinguli, die zowel door Gray als door Pribram en McGuinness tot het systeem wordt gerekend. Het is dan ook niet toevallig dat de hippocampus en de gyrus cinguli de twee belangrijkste structuren zijn van het circuit van Papez, dat volgens Papez (1937) verantwoordelijk is voor de emotieverschijnselen. Stimulatie van de gyrus cinguli veroorzaakt in ieder geval tekenen van emotionele opwinding: veranderingen in de hartslag, pupilver-wijding en pilo-erectie (Smith 1944). Beschadiging heeft ‘makheid’ tot gevolg, en verminderde emotionele responsiviteit. Er treedt bijvoorbeeld geen agressieve reactie meer op na frustratie (Brutkowski & Mempel 1961). Apen met beschadigingen in dit gebied houden zich niet meer bezig met het vlooien van hun makkers en tonen geen genegenheid meer voor ze. Ze behandelen mensen en andere apen als levenloze voorwerpen, gaan bovenop ze zitten of lopen tegen ze op. Ze vechten of vluchten niet wanneer ze daartoe gelegenheid krijgen. Over het algemeen gedragen ze zich alsof ze de emotionele consequenties van hun eigen handelingen niet kunnen overzien (Smith 1944; Ward 1948).
Laesies van de gyrus cinguli hebben als resultaat dat de drang om iets aan onaangename gebeurtenissen te doen weg lijkt te vallen, en dat de respons op signalen van naderende stressoren zwakker wordt (Gollender 1967). Bij mensen wordt cingulectomie toegepast in gevallen van ernstige, onbehandelbare pijn, bij ernstige angsttoestanden, dwanggedrag en driftaanvallen (Le Beau 1952; Powell 1979). De patiënten voelen de pijn nog wel maar hebben er niet al te veel last meer van. Wellicht heeft cingulectomie dit
| |
| |
effect omdat er een ingreep plaatsvindt in het systeem dat voorspellingen genereert omtrent externe gebeurtenissen en omtrent de consequenties van de handelingen van het subject zelf. Deze voorspellingen zijn waarschijnlijk gebaseerd op informatie die de gyrus cinguli krijgt aangevoerd van andere corticale centra (Gray 1982).
| |
7.6 ‘Gevoel’ en stemming
Zoals gezegd kan elektrische stimulatie van gebieden in de hypothalamus vormen van appetitief gedrag veroorzaken, zoals eten, drinken of copuleren. Het ligt voor de hand dat dit soort stimulatie aangenaam is voor de dieren, aangezien de lokaties vanwaar dit gedrag wordt opgewekt gewoonlijk ook de punten zijn vanwaar intracraniële zelfstimulatie (icss, ‘intracranial self-stimulation’) kan worden opgewekt, icss verwijst naar het verschijnsel dat een dier, wanneer het daar de gelegenheid toe krijgt, leert een pedaal in te drukken dat stroom aanzet in zijn hersenen. Het dier doet dit met duidelijke gretigheid, vaak in hoog tempo en gedurende lange tijd (Olds 1962). De ‘belonende’ punten bevinden zich verspreid over de middenhersenen en het limbische systeem. De zelfstimulatie is het sterkst in de laterale hypothalamus, in of bij de ‘medial forebrain bundle’ (mfg). De effecten die worden opgewekt kunnen verschillend van aard zijn. Door zelfstimulatie in de hypothalamus versnelt de hartslag; stimulatie van het septum lijkt een kalmerend effect te hebben, aangezien de hartslagfrequentie en de bloeddruk erdoor dalen (Routtenberg 1968). Of zelfstimulatie altijd het resultaat is van het aangename gevoel dat erdoor wordt opgewekt is niet zonder meer duidelijk. Op sommige punten lijkt het alsof de neiging tot zelfstimulatie en aversie tegen de stimulatie elkaar afwisselen. Bij mensen leidt hersenstimulatie, met name op bepaalde punten in de hypothalamus, tot zeer hevige aangename gevoelens, soms van orgastische aard (Sem-Jacobsen & Styri 1975; Bechtereva 1969; Heath 1976). In sommige gevallen blijkt het mogelijk om door stimulatie hevige pijn te verlichten (Heath 1976). Hierbij dient echter wel
de kanttekening geplaatst te worden dat bij mensen alle waarnemingen met betrekking tot de bevredigende effecten van hersenstimulatie afkomstig zijn van psychotische patiënten of patiënten bij wie, zoals gezegd, stimulatie pijnstillend werkt (Valenstein 1973). Desalniettemin bevestigt dit soort observaties de conclusie dat ‘gevoel’ een volledig centrale oorsprong kan hebben, zonder enige perifere of cognitieve inbreng.
De stimulatie van punten in de hypothalamus of andere structuren die afweergedrag opwekken wanneer er geschikte objecten aanwezig zijn, is duidelijk onaangenaam voor het dier. Het blijkt bijzonder gemakkelijk dieren een reactie aan te leren waardoor de stroom wordt uitgeschakeld
| |
| |
(Nakao 1958). Ze zijn tevens zeer vlot in het aanleren van vluchtgedrag en passief vermijdingsgedrag (Nakao 1967; Cox 1967). Bij mensen veroorzaakt de stimulatie van bepaalde punten (bijvoorbeeld de amygdala; Heath, Monroe & Mickle 1955; Chapman 1958) angstaanvallen. Bij patiënten met epileptische haarden en irriterende laesies in de temporaalkwab komen toestanden van hevige angst en van extatisch genot voor (Williams 1969).
Stemming en ‘gevoel’ zijn waarschijnlijk afhankelijk van de activiteit van de hersenmonoaminen noradrenaline, dopamine en/of serotonine. Er lijkt een verband te bestaan tussen de subcorticale catecholaminebanen en de eerder genoemde zelfstimulatiepunten, welke geconcentreerd zijn langs de mfb (zie Rolls 1975). De locus coeruleus, de plaats waar de dorsale noradrenerge bundel ontspringt, is een van de meest geschikte punten voor zelfstimulatie (Crow, Spear & Arbuthnot 1972). In hoeverre deze correlatie werkelijk opgaat staat echter nog ter discussie (Rolls 1975). De mate van zelfstimulatie kan worden beïnvloed door manipulatie van de noradrenaline- en dopamineneuronen, en van het metabolisme van deze neurotransmitters. De relaties blijken ingewikkeld en onduidelijk te zijn (Routtenberg 1978).
Over de exacte functie van de monoaminen bij de belonende effecten bestaat verschil van mening. Stein (1968) achtte de dorsale noradrenerge bundel verantwoordelijk voor het ‘bewerkstelligen van de gedragseffecten van beloning’, dat wil zeggen, van aangename signalen. Waarschijnlijk is dit onjuist, vernietiging van deze bundel heeft namelijk niet tot gevolg dat leerprocessen met voedsel, drinken of seks als beloning uitgeschakeld worden (zie Rolls 1975). Het is waarschijnlijker dat de monoaminen in verband staan met de aversieve beleving: stress veroorzaakt een toename in de omzet van noradrenaline (en andere monoaminen) in de hersenen (Stone 1983; Stein, Wise & Belluzi 1975). Gray trok op grond van een uitvoerig overzicht van alle gegevens de conclusie dat de functie van de dorsale noradrenerge bundel de overdracht van relevantiesignalen in het algemeen is, dat wil zeggen, het signaleren van gebeurtenissen die om verschillende redenen (beloning, straf, onbekendheid) van belang kunnen zijn.
Dopamine speelt duidelijk een rol bij de effecten van beloningen. Wanneer een dier dopamine-antagonisten krijgt toegediend verdwijnt het effect van elementaire beloningen op het leren en op zelfstimulatie (Fouriezos & Wise 1976). Het zou echter kunnen dat dopamineremming niet zozeer de hedonische beleving beïnvloedt, als wel de aandacht voor de stimuli. Dit blijkt bijvoorbeeld uit het feit dat door vernietiging van de dopaminebanen het subject minder gevoelig wordt voor diverse soorten zintuiglijke prikkels (Ljundberg & Ungerstedt 1976).
Serotonine speelt waarschijnlijk een rol bij de overdracht van signalen die te maken hebben met onaangename ervaringen. Stimulatie van de sero- | |
| |
toninebanen of van de serotonineomzet wekt angst op (Stein, Wise & Belluzi 1975). Door vernietiging van de banen en door toediening van antagonisten vermindert de responsonderdrukking die gewoonlijk onder invloed van straf optreedt. Benzodiazepinen neutraliseren het effect van stimulatie (zie Gray 1982, hoofdstuk 11 voor een bespreking). Op grond van deze gegevens veronderstelt Gray dat de serotoninebanen de signalen van onaangename gebeurtenissen overbrengen. Volgens Pribram en McGuinness (1975) zijn de serotoninebanen verantwoordelijk voor de overdracht van relevantiesignalen, alsook voor gedragsinterruptie en aandachtsactiviteit in het algemeen.
Het is waarschijnlijk dat de hersenmonoaminen een rol spelen in het ontstaan van depressieve stemmingen en misschien ook depressieve psychosen. Hun omzet neemt toe onder invloed van stress. Men heeft wel beweerd dat langdurige stress leidt tot een gebrek aan monoaminen in de hersenen; volgens sommigen gaat het hier in het bijzonder om noradrenaline (Weiss, Glazer & Poherecky 1976), volgens anderen om alle drie de monoaminen (Anisman & Zacharko 1982). Er zijn aanwijzingen dat dit vooral of zelfs uitsluitend voorkomt wanneer het subject geen invloed kan uitoefenen op de stress en alle hanteringsmechanismen ontoereikend blijken, zoals bij het aangeleerde hulpeloosheidsparadigma (Weiss e.a. 1976; Anisman & Zacharko 1982; Laborit 1979). Depressie zou derhalve het resultaat kunnen zijn van uitputting van een of meerdere monaminen: noradrenaline (Schildkraut & Kety 1967), serotonine (Van Praag 1983), of een combinatie met onder andere adrenaline (Roth 1983). Een andere mogelijkheid is dat depressie wordt veroorzaakt door neurale overstimulatie ten gevolge van de toegenomen neurotransmitteromzet (Stone 1983). De relatie tussen depressieve stemmingen en psychosen en monoamine-uitputting of verhevigde monoamineomzet is een controversiële kwestie die voor een groot deel op indirecte gegevens berust. Dat er een relatie bestaat tussen depressieve toestanden - en stemming in het algemeen - en de monoaminen is echter wel waarschijnlijk; het blijkt bijvoorbeeld uit het effect van stemmingsveranderende middelen op het monoaminemetabolisme of de monoamineoverdracht. Amfetamine, met zijn activerende en euforiserende werking, is een catecholamine-agonist. Tricyclische antidepressiva zoals imipramine remmen de monoamineheropname. Monoamineoxidaseremmers, stoffen
waardoor de impulsoverdracht door middel van monoaminen wordt bevorderd, hebben een antidepressieve werking, terwijl zowel amfetamine als de monoamineoxidaseremmers zelfstimulatie bevorderen. Middelen waardoor het monoaminemetabolisme wordt verstoord, zoals reserpine, kunnen ernstige depressieve toestanden veroorzaken.
Van groot belang bij de processen die aan ‘gevoel’ en stemming ten grondslag liggen zijn de endorfinen. De endorfinen zijn opiumachtige stoffen voorkomend in de hersenen en elders in het lichaam; ze worden vooral
| |
| |
tijdens stress afgescheiden. Ze hebben sterke affectieve effecten. Bepaalde endorfinen produceren net als morfine aangename gevoelens en euforie; ratten leren zelfstimulatie aan voor met- en leu-encefaline (Belluzi & Stein 1977), en deze stoffen lijken, net als morfine, verslavend te zijn. Toediening van bèta-endorfine blokkeert de gevoeligheid voor pijn (de pijn is er wel, maar wordt niet als hinderlijk ervaren) (Snyder & Childers 1979), en vermindert door pijn opgewekte reflexen (Belluzi e.a. 1976). Door de bètaendorfine-antagonist naloxon worden deze reflexen juist versterkt (Fanselow & Bolles 1979). Andere endorfinen hebben een slaapverwekkend, kalmerend effect: gamma-endorfine en daarvan afgeleide stoffen versnellen de extinctie van geconditioneerd vermijdingsgedrag (De Wied e.a. 1978). Het derivaat dt-gamma-g zet, net als haloperidol, proefdieren aan tot het vastgrijpen van het traliewerk van hun kooi. Verzwakking van de aversieve beleving zou de oorzaak kunnen zijn van deze effecten. Op grond hiervan formuleerde Wise (1982) zijn ‘anhedonie-hypothese’ ter verklaring van het effect van de narcoleptica. Deze stoffen zouden de via de dopaminebanen overgebrachte negatieve hedonische toon kunnen verminderen. De hypothese zou dus ook op gamma-endorfine van toepassing kunnen zijn.
Het onderzoek naar de emotionele implicaties van natuurlijk voorkomende endorfinen is nog nauwelijks begonnen. Het valt te verwachten dat deze implicaties niet gering zullen blijken, vooral gezien het feit dat de endorfinen niet zozeer als neurotransmitters functioneren, maar als neuromodulators.
Behalve neurotransmitters en endorfinen zijn er nog andere neurohumorale stoffen die invloed uitoefenen op het emotionele gedrag en de emotionele beleving. De belangrijkste is het adrenocorticotroop hormoon, acth. acth is wel ‘het vrees-mediërende neurohormoon’ genoemd (Jacquet 1980). Het is een stof die de vreesreactie beïnvloedt. De verwerving en de extinctie van zowel actief als passief vermijdingsgedrag nemen af na verwijdering van het voorste deel van de hypofyse, waardoor de acth-produktie wegvalt. Wanneer vervolgens acth wordt toegediend herstelt zich het vermogen vermijdingsgedrag aan te leren (De Wied 1964; Levine & Jones 1965). acth-4-10-implantaten in het mesencefalon en diëncefalon kunnen het aanleren van vermijdingsgedrag versterken (De Wied e.a. 1978; zie De Wied 1980). De extinctie van appetitief gedrag (prestatie voor een voedselof seksuele beloning) neemt eveneens af wanneer de acth-produktie wordt geblokkeerd en wordt weer normaal zodra het acth-niveau zich herstelt. Deze effecten lijken centraal te worden voortgebracht, zoals blijkt uit het feit dat toediening van acth invloed heeft op de elektrische activiteit van de hippocampus (Urban & De Wied 1976).
Andere hormonen met een centrale activiteit zijn de corticosteroïden en testosteron, en in feite alle hormonen die deel hebben aan het hypothalamus-hypofyse-gonadale systeem. De activatie van dit systeem, hetzij door
| |
| |
directe hormonale manipulatie zoals het toedienen van testosteron (Bermond 1978), hetzij op andere wijze, bevordert zowel seksueel als agressief gedrag, en vermoedelijk ook de daarmee gepaard gaande emoties van seksuele opwinding en woede of ergernis (Beach & Holtz 1946; Bermond 1982; Poshilavov 1982). Ook het premenstruele syndroom is een duidelijke aanwijzing dat de drempel voor irritatie, woede en depressiviteit onder invloed van hormonen kan worden verlaagd (Parlee 1973; Smith 1975).
De corticosteroïden en acth hebben uiteenlopende effecten op vermijdingsgedrag, vrees en agressie. De uitwerking van deze stoffen op het gedrag is wisselend en complex (zie Leshner 1978,1983 voor een bespreking). Als algemene verklaring kan men misschien het beste uitgaan van de hypothese dat deze stoffen de aandacht en de intentionele paraatheid van het subject versterken, en daarmee de emotionele impact van de situatie (Bohus 1975; Leshner 1983). Ze oefenen dus wel invloed uit op de stemming, maar zouden dat dan niet op een directe en specifieke manier doen. Ze kunnen de stemming bijvoorbeeld beïnvloeden door de gevoeligheid voor stimuli uit de omgeving te verhogen, en de bereidheid van het subject om op die stimuli te reageren. De complexiteit van de dispositionele antecedenten van de emotie wordt hierdoor eens te meer benadrukt.
De interpretatieproblematiek die uit het bovenstaande naar voren komt wijst er verder op dat de specifieke werking van de meeste neurohumorale factoren op stemming en ‘gevoel’ onduidelijk is, en op verschillende manieren geïnterpreteerd kan worden. Dat wil zeggen, het is onduidelijk welk proces ze precies beïnvloeden, en welk proces derhalve verantwoordelijk is voor de effecten op de emotionele beleving. Is het effect direct, dat wil zeggen brengen de monoaminen ‘gevoel’ teweeg, of zijn ze van invloed op de activatie in het algemeen, of op de specifieke reactieneiging van het subject, en daarmee op diens competentiebesef? Of oefenen ze invloed uit op de alertheid van het subject, en daarmee op diens gevoeligheid voor prikkels? Een van de weinige analyses die een eind in de richting komen wat betreft de verklaring van de emotionele beleving is de angsthypothese van Gray. Deze schrijft namelijk aan de activiteit van het neurofysiologische systeem in kwestie, het gedragsinhibitiesysteem, precies die cognitieve en hedonische antecedenten en gedragsmatige consequenties toe op grond waarvan die emotionele beleving te verklaren is. Het hedonische karakter van de antecedenten blijft echter onverklaard en is misschien ook niet verder herleidbaar.
| |
7.7 Verschillen tussen de hersenhelften
Er wordt tegenwoordig betoogd dat de twee hersenhelften wat betreft de emotie niet precies gelijkwaardig zijn. De rechter hersenhelft is, bij rechts- | |
| |
handigen althans, mogelijk emotioneler dan de linker.
Er zijn enige aanwijzingen die deze hypothese ondersteunen. Een overzicht van die gegevens is te vinden bij Flor-Henry (1979), Campbell (1982) en Kinsbourne en Bemporad (1984). Hieruit blijkt bijvoorbeeld dat depressieve patiënten aan de linkerhand een sterkere elektrodermale respons vertonen dan aan de rechter. Epileptische haarden in de rechter temporaalkwab veroorzaken meer stemmingsstoornissen dan haarden links. Pijn na amputaties wordt links erger gevoeld dan rechts (vergeet niet dat zowel de gevoels- als de motorische zenuwbanen gekruist verlopen). De gegevens zijn zwak. Sterker zijn de gegevens omtrent de verschillen tussen de hersenhelften bij normale individuen. Er is bijvoorbeeld onderzoek gedaan naar de gevoeligheid voor emotionele stimuli, zoals de affectieve waarde van de intonatie van de stem (bijv. Safer & Leventhal 1977) of van de gelaatsexpressies van anderen. In elf onderzoeken bleek de rechter hersenhelft wat betreft dit laatste superieur aan de linker (Campbell 1982). De rechter hersenhelft is eveneens beter in een groot aantal nonverbale taken. In minstens een van deze onderzoeken bleek de superioriteit van de rechter hersenhelft in de verwerking van emotionele informatie (McKeever & Dixon 1981). Experimenten met patiënten met laesies in de rechter of linker hersenhelft hebben tegenstrijdige resultaten opgeleverd, ofschoon over het algemeen wel is gebleken dat letsel in de rechter hersenhelft een verstorender effect op nonverbale emotieherkenningstaken had dan letsel in de linker hersenhelft.
Gelaatsexpressies zijn doorgaans meer geprononceerd op de linker gezichtshelft dan op de rechter, hetgeen erop zou kunnen duiden dat de rechter hersenhelft meer bij de gelaatsexpressie is betrokken dan de linker (Sackheim, Gur & Saney 1978). Dit lijkt evenwel meer te gelden voor mimiek dan voor echte gelaatsexpressies (Ekman, Hager & Friesen 1981). Voor zover het hier gaat om de gelaatsexpressie als zodanig - en niet om een grotere mobiliteit van de linker gezichtshelft (Campbell 1982) - duidt dit verschijnsel waarschijnlijk eerder op een sterkere inhibitie van de linker hersenhelft dan op emotionaliteit van de rechter: de echte gelaatsexpressies hebben een subcorticale oorsprong (Rinn 1984).
Een andere veronderstelling is dat er een kwalitatief emotioneel verschil bestaat tussen beide hersenhelften. Hiervoor bestaan nogal wat aanwijzingen, waarvan wederom overzichten zijn gegeven door Flor-Henry (1979), Campbell (1982) en Kinsbourne en Bemporad (1984). De rechter hersenhelft lijkt meer geneigd tot droevige of depressieve stemmingen, de linker tot euforie. Het lijkt er bijvoorbeeld op dat elektroshocks rechts effectiever zijn voor het opheffen van depressieve toestanden dan links (Galin 1974). Hersenletsel aan de linkerkant brengt droevige stemmingen teweeg, terwijl het rechts in veel gevallen leidt tot emotionele vlakheid of joligheid (Gianotti 1972). De interpretatie van de resultaten is echter een complexe aan- | |
| |
gelegenheid, en men heeft de resultaten bovendien niet altijd kunnen bevestigen door middel van herhaald onderzoek (Campbell 1982). Desondanks zijn ze suggestief en wijzen ze over het algemeen in dezelfde richting (zie ook Fox & Davidson 1984). Een hypothese die het vermelden waard is, is die van Kinsbourne en Bemporad (1984). Volgens deze onderzoekers staat het linker frontale gebied in dienst van de gedragsbesturing, of ‘activatie’ in de betekenis van Pribram en McGuinness. Letsel in dit gebied leidt derhalve tot apathie en onvermogen tot gedragsplanning, met depressiviteit als secundaire consequentie. Het rechter frontale gebied regelt de besturing van emotie, oftewel ‘arousal’, en de evaluatie van emotioneel relevante informatie (zie ook Tucker 1981). Letsel in dit gebied heeft onaangepaste en slecht gecontroleerde emotionele reacties of onverschilligheid tot gevolg. |
|
