|
| |
| |
| |
De nachtelijke activiteit van de hersenen: slaap en droom
A. Bauchau
Wat weten wij fysiologisch van slaap en droom? Twee theorieën die eens furore maakten, maar nu voorgoed voorbijgestreefd zijn, kunnen ons op weg helpen in de doolhof van het probleem.
De eerste stelde dat het organisme in de loop van de dag een niet nader bepaalde toxine (giftige stof) accumuleerde die werkte als een narcoticum en tenslotte de slaap veroorzaakte; in de slaap werd die toxine dan weer geëlimineerd. Vooral het onderzoek van Siamese tweelingen heeft deze theorie ontzenuwd. Indien de slaap veroorzaakt wordt door een toxine, dan zouden Siamese tweelingen, wier bloedsomloop in elkaar vertakt is, tegelijk moeten inslapen en wakker worden. Welnu, dat is helemaal niet het geval, zoals is gebleken uit de observatie van een Russische ‘psodyme’ tweeling.
Volgens de tweede theorie ontstond de slaap uit een tijdelijke vermindering van de bloedstroom door de hersenen. De zenuwcellen zouden minder zuurstof krijgen, ze zouden lijden aan een goedaardige verstikking, waardoor hun activiteit wordt stopgezet of geremd. Proefondervindelijk is echter gebleken dat de bloedstroom door de hersenen tijdens de slaap niet afneemt, maar integendeel kennelijk, zij het lichtjes, toeneemt. Het zuurstofverbruik blijft praktisch even hoog als in wakende toestand.
Chemisch zijn de hersenen dus even actief wanneer ze slapen als wanneer ze over al hun psychische vermogens beschikken. Deze paradox betekent natuurlijk niet dat de topografie van die activiteit in beide gevallen identiek is.
| |
Elektrische studie van de slapende hersenen
De eerste aanzet voor een fysiologische verklaring van de slaap dateert van na de eerste wereldoorlog. Om de elektrische stroom die door de hersenen loopt te detecteren, ontwerpt Hans Berger, een psychiater uit Jena, een apparatuur die voor die tijd bijzonder ingewikkeld was. In 1924 slaagt hij erin het eerste encefalogram op te nemen. Scrupuleus, bang of bedeesd,
| |
| |
publiceert hij het resultaat pas vijf jaar later, in 1929. Reactie in wetenschappelijke kringen: bitter weinig, geen mens neemt de moeite Bergers proefnemingen over te doen. Die onverschilligheid is wel niet goed te praten, wel te verklaren: het ongeordende tracé van Bergers encefalogram, resultante van de globale activiteit van miljarden cellen, schijnt nu niet direct een ideale grondslag te bieden voor een analyse van zo complexe processen als die van de hersenen. Bergers opnemingen zien er trouwens helemaal niet spectaculair uit: gewone elektrische oscillaties met kleine amplitude, een tiental per seconde. Bovendien, Berger was een psychiater, en hebben psychiaters niet de naam dat zij altijd wel argumenten vinden voor de meest buitenissige ideeën als zij zelf maar overtuigd zijn van de juistheid ervan? Zo kon de grote Engelse fysioloog Adrian in 1934 nog verklaren: al die oscillaties lijken mij ‘hopeloos constant’.
Maar het zou niet lang duren voor Bergers apparaat geperfectioneerd werd en de ervaring een meer verantwoorde interpretatie van zijn encefalogrammen mogelijk maakte. Alles wat wij op het ogenblik weten over slaap en droom, danken wij aan Bergers ontdekking. Met name denken wij dan aan het prachtige werk van de ploeg van Kleitman aan de Universiteit te Chicago, met Aserinsky, Dement en Wolfert; aan het meer recente werk ook van een team in Lyon: Jouvet, Klein en Ruckebusch. Vindingrijk en volhardend hebben zij Bergers vondst benut om het onbekende land van de slapende hersenen in kaart te brengen.
De elektrische activiteit in de verschillende gebieden van de hersenschors of van de spieren aan de oppervlakte van het lichaam is tegenwoordig heel gemakkelijk te registreren. Op ieder gewenst punt van schedel, oogkassen, borst of rug kun je een elektrode bevestigen en deze verbinden met een registreerapparaat. Proefpersonen, die een nachtje in het laboratorium willen komen slapen, zijn blijkbaar gemakkelijk te vinden (bij Klein kregen ze vijf dollar per nacht). Zo kan het verloop van de zenuw- en spieractiviteit tijdens slaap en droom objectief geobserveerd worden.
Figuur 1 toont hoe zo'n registratie er uit ziet. In waaktoestand vertonen de hersenen heel snelle en onregelmatige oscillaties, 8 tot 13 cycles per seconde. Dit is het zogenaamde ‘alfa-ritme’. Bij het inslapen vertraagt dit ritme (4 tot 6 cycles per seconde, in fase 1). Daarop volgen oscillaties die gegroepeerd zijn in bundels met variërende amplitude (14 tot 16 cycles per seconde, in fase 2). In fase 3 sterft het ritme langzaam weg. Tenslotte, in fase 4, treden de delta-golven op met grote amplitude (2 tot 4 cycles per seconde). Waarschijnlijk wijzen deze op een toenemende synchronisatie van de celactiviteiten van de schors. Het subject slaapt nu heel diep en rustig. De ademhaling gaat langzaam, het hart klopt trager, de oogballen maken trage horizontale bewegingen. Al deze verschijnselen wijzen op wat
| |
| |
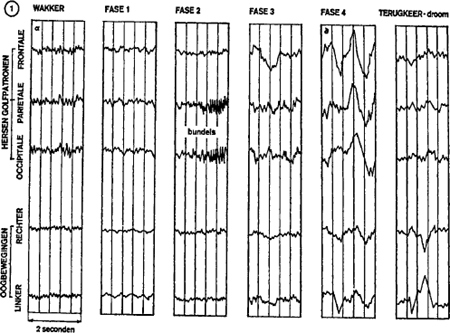
genoemd wordt de ‘trage slaap’. De drempel van het wakker worden gaat steeds hoger liggen. Maar nu volgt een terugkeer naar fase 1, en meteen opnieuw dezelfde cyclus. In de loop van een nacht doorlopen wij aldus verschillende van die cycli. Iedere cyclus duurt ongeveer 90 minuten. Tegen het einde van de nacht wordt de slaap wel minder vast: fase 4, de diepe slaap, wordt niet meer bereikt, we springen vaak van fase 2 of 3 direct terug over naar fase 1.
Drie tot vijf keer per nacht, op het ogenblik van de terugkeer naar periode 1, wordt het rustige tracé van het encefalogram ineens heftiger: het vertoont gelijkenis met dat van hersenen in waaktoestand. De oogbollen gaan bewegen, alsof ze een boeiend schouwspel volgden; ademhaling en hart gaan sneller. De tonus van de halsspieren, die in de trage slaap gehandhaafd blijft, verdwijnt; de slaper blijft doorgaans onbeweeglijk. De drempel van het wakker worden ligt nu heel hoog. Wordt hij overschreden en wordt de slaper dus abrupt wakker gemaakt, dan herinnert hij zich in 80% van de gevallen dat hij gedroomd heeft en kan hij gemakkelijk beschrijven wat hij ‘zag’.
Figuur 2 synthetiseert de observaties waargenomen bij eenzelfde individu gedurende 13 opeenvolgende nachten. De horizontale staafjes duiden begin en duur van de oogbewegingen aan. Het cyclische karakter van het fenomeen is duidelijk te zien. Normaal bewegen de ogen zich in een horizontaal vlak. Bewegen ze zich in een verticaal vlak, dan verklaart het subject,
| |
| |
wanneer het wakker gemaakt
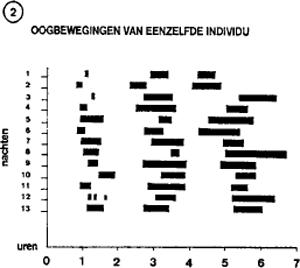
wordt, dat het iets waarnam dat zich van boven naar beneden verplaatste. Verraadt het encefalogram weinig of geen oogbewegingen, dan verklaart de proefpersoon dat hij iets zag dat ver af lag. Er bestaat dus een bevredigende correlatie tussen de bewegingen van de ogen en de aard van het droombeeld. Overigens, hoe actiever het subject in zijn droom betrokken is, hoe meer de ogen bewegen.
Beweging van de ogen en verdwijning van de tonus van de voorste halsspieren zijn de zekerste criteria om te zeggen: hier wordt gedroomd.
In figuur 3 zien we het normale verloop van de slaap: opeenvolgende cycli van ongeveer 90 minuten; maximale diepte van de slaap in het begin, daarna steeds oppervlakkiger slaap naarmate de nacht vordert; heftige
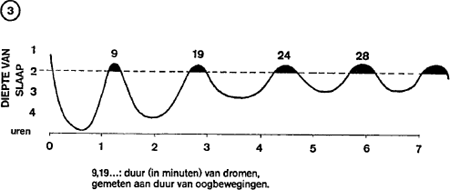
oogbewegingen op het ogenblik dat de slaper uit de diepe slaap terugkeert naar fase 1; ontwaken tenslotte, dat voorafgegaan wordt door een laatste droom, de droom die doorgaans het gemakkelijkst in de herinnering blijft. Hieruit blijkt dat de droom niet een flitsend visioen is, dat in enkele seconden tijd een indrukwekkend aantal wederwaardigheden opeenhoopt, maar een proces dat ongeveer in hetzelfde tempo verloopt als in het werkelijke leven. Dement heeft dit overtuigend aangetoond. Hij laat een van zijn proefpersonen tien minuten dromen, zoals dat af te lezen is van de snelle oogbewegingen; dan sluipt hij de kamer binnen en spuit wat water op de rug van de slaper; hij wacht 30 seconden, maakt hem abrupt wakker en begint hem te ondervragen. De proefpersoon begint een toneelstuk te
| |
| |
beschrijven, waarin hijzelf een rol speelt: ‘Ineens valt de hoofdactrice neer. Water stroomt over haar neer. Ik loop naar haar toe, voel op mijn beurt water over hoofd en rug lopen. Waarom is zij gevallen? Ik zie kalkpuin op haar kleren, ik kijk omhoog en zie een gat in het plafond. Ik trek haar in de coulissen en roep dat ze het doek neer moeten laten. Dan ben ik wakker geworden’. Dement laat hem daarop heel dit verhaal naspelen, vanaf het ogenblik dat hij dat water heeft gewaargeworden. De reconstructie vraagt nauwelijks minder dan 30 seconden, ongeveer de tijd dat Dement gewacht heeft.
Steunend op vele analoge gegevens, meent Dement dat de duur van onze dromen varieert van 8 tot 30 minuten, soms meer. De eerste droom van de nacht schijnt over het algemeen korter te zijn (gemiddeld 10 minuten in 71 geobserveerde nachten), de tweede en de volgende duren van 20 tot 35 minuten. Wij besteden dus een of twee uur van onze slaap aan de droom, d.i. 18 à 25% van de nacht.
Dit geldt voor volwassenen. De hersenactiviteit van de foetus daarentegen lijkt nogal anarchistisch. Pas vanaf de 6e maand wordt het encefalogram wat regelmatiger en vertoont het een cyclisch karakter. In de 8e maand wordt het onderscheid tussen slaap en waaktoestand goed zichtbaar. De cycli zijn bij pasgeborenen en kinderen korter (Aserinsky): slechts een uur, waarna de baby wakker wordt, even woelt, en een nieuwe periode van 60 minuten begint. De cycli worden geleidelijk langer, tot ze gelijk worden aan die van volwassenen. Hetzelfde zien we trouwens in de hart-, ademhalings- en maagcyclus, hetgeen erop schijnt te wijzen dat aan alle samen een fysiologische basiscyclus ten grondslag ligt.
Concluderend: in één slaapperiode zijn duidelijk twee toestanden te onderscheiden: de langzame slaap en de snelle of onirische slaap, ook de paradoxale slaap genoemd. De globale activiteit van de hersenen vermindert blijkbaar niet; ze is alleen anders, op een andere basis georganiseerd dan wanneer wij wakker zijn.
Deze onderzoekingen hebben een eind gemaakt aan tal van legenden, zelfs van vaste overtuigingen. Hoeveel mensen zijn er niet die beweren dat zij nooit of zelden dromen? Welnu, alle proefpersonen van Kleitman die dachten dat zij nooit droomden, erkenden dat zij bezig waren te dromen op het ogenblik dat zij wakker werden gemaakt. Het lijkt dus wel heel waarschijnlijk, zoniet zeker, te zijn dat wij allemaal iedere nacht dromen en iedere nacht meer dan eens. Maar heel weinig herinneren wij ons daarvan. Alleen als we wakker worden uit een droom die nog aan de gang was of net afgelopen, kunnen wij er ons gemakkelijk iets van herinneren. En hoe langer de droom duurt over de paradoxale fase heen, des te gemakkelijker verdwijnt hij voorgoed uit het geheugen.
| |
| |
| |
Slaap en droom bij dieren
Willen we deze fenomenen verder bestuderen, dan moeten we gaan experimenteren. Dat is niet zo makkelijk en niet altijd onschuldig: opzettelijk gaan we het proces verstoren en brengen we er traumatismen in aan. Dat kunnen we alleen doen met dieren. Maar dan rijst eerst de vraag: slapen dieren echt? En vooral: dromen ze?
Het best gaan we te rade bij de soorten die het dichtst bij ons staan.
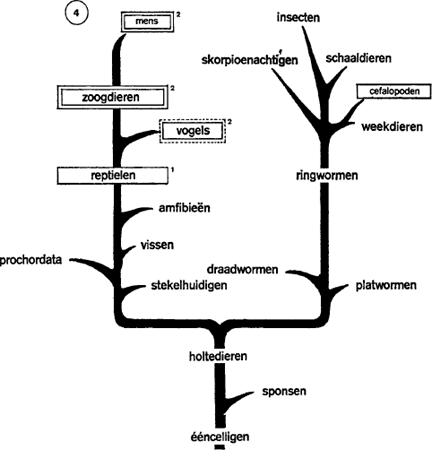
FILOGENETISCHE VERSPREIDING VAN ‘TRAGE SLAAP’ 1 en ‘PARADOXALE SLAAP’2
Figuur 4. Bij insecten, schaaldieren, gasteropode weekdieren (slakachtigen) zijn perioden van inactiviteit, zelfs van schijnbare rust waar te nemen, maar van echte slaap kun je hier niet spreken. Cefalopode weekdieren daarentegen (zoals de inktvis) kennen een sterke cerebrale concentratie en een dagelijks gedrag dat heel dicht bij de slaap staat: onbeweeglijkheid,
| |
| |
daling van de prikkelbaarheid, inkrimping van de pupillen, vertraging van de ademhaling, enz.. Voorlopig zijn dit de enige ongewervelde dieren die deze fenomenen vertonen.
Onder de gewervelde dieren kennen de nachtvissen (zeepaling en pieterman) blijkbaar geen slaap. Bij andere wordt de activiteit overdag afgewisseld met inactiviteit 's nachts. Maar slapen ze daarom? Dat is niet zo zeker. Bij de amfibieën wordt het al iets duidelijker. Sommige tritons (watersalamanders) bijvoorbeeld komen 's nachts uit het water, sluiten de ogen, worden zintuiglijk en motorisch minder prikkelbaar, en hernemen hun wakkere activiteit pas tegen zonsopgang.
Bij de reptielen is er geen twijfel meer: slangen, hagedissen, krokodillen, schildpadden slapen overvloedig. In 1963 heeft Klein, van de school van Lyon, een encefalogram geregistreerd van inslapende schildpadden. Hij constateerde dezelfde symptomen als in de trage slaap van de mens: merkelijke ontspanning van de halsspieren, vertraging van hart en ademhaling; er is geen reactie meer op audio-visuele prikkels en tamelijk sterke huidprikkels zijn nodig opdat het dier kop en poten onder het schild terug zou trekken. Snelle slaap of droom kon men echter niet ontdekken. De schildpad besteedt de helft van haar lange bestaan aan de slaap, dromen echter blijven haar blijkbaar ontzegd.
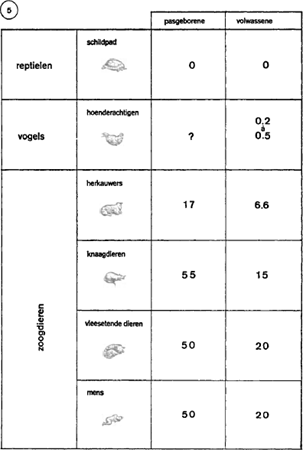
Klein heeft ook onderzocht hoe kippen en duiven inslapen. Ze trekken de nek in, leggen de kop vast op de krop, sluiten de oogleden en ontspannen de tonus van de spieren. Maar nu manifesteert zich iets nieuws, zij het nog heel bescheiden: na een half uur trage slaap verschijnen op het encefalogram paradoxale fasen, die gepaard gaan met enkele snelle, ongeordende oogbewegingen. Dit schijnt op vluchtige dromen te wijzen, die niet langer duren dan 5 of 6 seconden, in totaal slechts 0.2 à 0.5% van de totale slaapduur (figuur 5).
Bij de zoogdieren tenslotte vinden we de verschillende manieren van slaap in hun volledige expressie. Echter niet bij alle zoogdieren in dezelfde graad. Varkens bijvoorbeeld zijn goede slapers, zij nemen zelfs een comfortabele siësta. Volwassen herkauwers daarentegen slapen veel minder: het schaap bijvoorbeeld 48 minuten in 24 uur, en dan is die slaap nog heel licht.
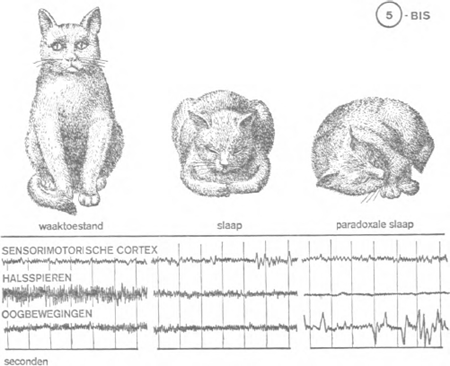
Gelukkig voor de fysiologen zijn de vleesetende dieren voortreffelijke slapers, en daaronder zijn er die heel dicht bij ons staan: de hond en de kat. De kat bijvoorbeeld besteedt 65% van haar tijd aan zalige slaap. En het verloop daarvan is nauwkeurig af te lezen van een encefalogram: doorgaans 10 à 20 minuten lichte slaap, te vergelijken met de trage slaap van de mens; daarna een paradoxale periode van 6 à 7 minuten, die schijnt te beantwoorden aan de droomfase in de menselijke slaap: totale ontspanning van de nekspieren, ongeordende bewegingen van de oogbollen. Deze op- | |
| |
DUUR VAN PARADOXALE SLAAP IN % VAN TOTALE DUUR VAN SLAAP
eenvolging wordt verschillende keren herhaald, met een tussenpauze van telkens 25 minuten. De kat levert aldus goed vergelijkingsmateriaal ( figuur 5 bis).
| |
Fysiologisch onderzoek van slaap en droom
Vanzelf rijst een eerste vraag: bezit het zenuwstelsel een bepaalde structuur die slaap veroorzaakt, of volgt slaap gewoon uit de vertraging of stilstand van de activiteit van een centrum dat verantwoordelijk is voor het wakker zijn?
| |
| |
| |
a. De substantia reticularis (het reticulair stelsel): waakzaamheidscentrum
Sinds 1949 is het bestaan bekend van een zenuwformatie die verantwoordelijk is voor het wakker zijn. De anatomie leert dat alle zintuigzenuwen die uitgaan van de periferie van het lichaam, wel uitlopen op de schors, maar onderweg zijn er vertakkingen van zenuwnetten; deze worden geleid naar een groep zenuwcellen, gelegen in het centrum van de hersenstam: de ‘substantia reticularis’ of het ‘reticulair stelsel’ (figuur 6).
Dit reticulair stelsel is niet groter dan een pink, maar vormt
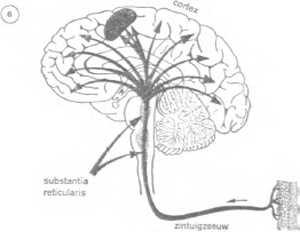
een heel interessante structuur. Ze werd onderzocht door de Amerikaan Mazoun en de Italiaan Moruzzi, aan de Northwestern University. Een zwakke elektrische stroom door het reticulair stelsel volstaat om een slapende kat wakker te maken; wordt het vernietigd, dan volgt een coma waaruit de kat nooit
| |
| |
meer wakker wordt. Kennelijk vormt het dus een waakzaamheidscentrum. Het is als een vooruitgeschoven wachtpost in het zenuwstelsel: het registreert alle perifere zintuigelijke boodschappen, en wanneer deze sterk of significatief genoeg zijn, maakt het de schors wakker en bereidt ze voor op hun komst. Komt het niet tussenbeide, dan bereiken de signalen wel hun bestemming, maar ze worden niet herkend, niet geïnterpreteerd en wekken dus geen enkele intelligente reactie. Deze functie verklaart bijvoorbeeld hoe een moeder in haar slaap het lawaai van de straat niet hoort, maar wakker wordt zogauw haar kind zich laat horen. Het huilen van het kind waarschuwt direct de substantia reticularis, waarvoor het een precieze betekenis heeft; anonieme geluiden daarentegen wekken geen enkele reactie.
Deze anatomische structuur verklaart ook de werking van verdovende middelen en slaapmiddelen: ze blokkeren de toegang tot de substantia reticularis, zonder de toegang van de zenuwimpulsen tot de schors ernstig te beïnvloeden. Opwekkende middelen daarentegen vergemakkelijken de toegang van de impulsen, brengen het reticulair stelsel in staat van alarm, en versterken aldus de waakzaamheid.
Verder regelt en verfijnt het reticulair stelsel enigszins de motorische reacties, vrijwillige zowel als reflexe. Een fraaie demonstratie daarvan vinden wij bijvoorbeeld in de studie van knieschijfreflexen. Van een gevoelloos gemaakte aap legt men een van de poten op een stang; aan een van de tenen maakt men een stilet vast; tikt men op de knieschijf, dan registreert het stilet de reflex op een cylinder. Laat men vooraf een zwakke elektrische stroom door de top van de substantia reticularis gaan, dan is de reflex sterker en omvangrijker; een prikkel onder in de substantia reticularis daarentegen maakt de motorische reactie zwakker.
Is hiermee alles verklaard? Is slaap dan niets anders dan de tijdelijke uitschakeling van die waak-rol welke de reticularis speelt ten opzichte van de schors? Volgens de Belgische fysioloog Bremer zou die uitschakeling dan bijvoorbeeld veroorzaakt kunnen worden door een verzwakking
van de uitwendige prikkels: wij slapen inderdaad gemakkelijker in als het stil en donker is.
| |
b. De raphé: slaapcentrum
Een nieuw experiment van Moruzzi heeft deze opvatting echter grondig gewijzigd. Om de wer- | |
| |
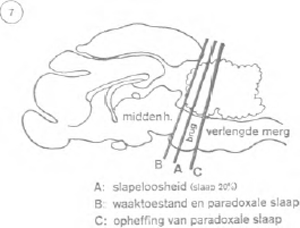
king van het reticulair stelsel verder te onderzoeken, voerde hij een sectie uit op de hersenstam van een kat, ongeveer midden in de brug van Varol, juist voor de plaats van uittreding van de drielingzenuw (figuur 7). Gevolg: de kat sliep nog slechts 20% van de dag, in plaats van 65%. Het alfa-ritme van de hersenschors en de oogbewegingen verrieden overigens een kennelijke waaktoestand. De operatie scheen het reticulair stelsel bevrijd te hebben van de remmende controle van bepaalde centra van de hersenstam. Wat voor centra zijn dat en waar liggen ze? Jouvet denkt aan een groep zenuwcellen, bekend onder de naam ‘raphé’ (gr. naad), die zich bevindt op de middellijn van het verlengde merg (figuur 8). Worden deze cellen voor 80% vernietigd, dan ontstaat er een nog duidelijker slapeloosheid dan in het experiment van Moruzzi: de kat slaapt nog slechts 10% van de tijd. Een radicalere vernietiging, 100%, is niet mogelijk: dat zou het dier met overleven.
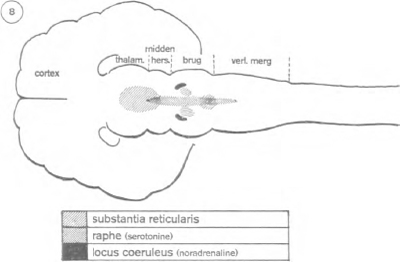
Later ontdekken twee Zweedse geleerden, Dahlström en Fuxe, dat de cellen van de raphé, onder ultraviolette stralen, een sterke gele fluorescentie vertonen. Dit betekent dat ze een opmerkelijke kwantiteit serotonine bevatten. Welnu, sinds lang weten we dat deze scheikundige stof (indol) een krachtig effect heeft op de hersenen, zoals overigens op andere organen. Het feit dat het ook aanwezig zou zijn op zo'n strategische plaats als de raphé, opent de weg naar een biochemische interpretatie van de slaap. Jouvet heeft inderdaad kunnen aantonen dat er een nauw parallellisme bestaat tussen de omvang van de beschadiging welke aan de raphé wordt
| |
| |
aangebracht, de vermindering van slaap en een verminderde concentratie van serotonine in de hersenen.
Voorlopig samenvattend kunnen we dus zeggen: het ontstaan van de slaap schijnt afhankelijk te zijn van het samenspel van tenminste twee formaties: het waakzaamheidscentrum in het reticulair stelsel en het slaapcentrum in de raphé. Serotonine kan dan de chemische tussenschakel zijn in de remmende werking van het tweede op het eerste.
Dit is echter slechts een voorlopig operationeel schema. De slaap is ook afhankelijk van een werking van de voorste hersencentra (thalamus en cortex). Worden een kat deze afgenomen, dan worden er geen golven meer geregistreerd die kenmerkend zijn voor de trage slaap. Maar de reden van deze interventie van de bovenste centra is nog niet duidelijk.
| |
c. De Locus Coeruleus: droomcentrum
Paradoxaal genoeg weten we op het ogenblik beter welke zenuwstructuren nodig zijn voor de droom dan voor de slaap. Een sectie op de hersenen van de kat juist voor de Brug van Varol supprimeert praktisch de werking van de voorste hersenen. Het dier blijft nog maanden leven, maar gaat ononderbroken, steeds weer opnieuw direct over van het wakker zijn naar de paradoxale slaap (cfr. figuur 7). Dezelfde operatie achter de Brug stelt nog meer zenuwcentra buiten werking en schakelt ook elke paradoxale fase definitief uit. Waarschijnlijk hangt deze dus af van bepaalde zenuwcellen van de Brug. Jouvet heeft aangetoond dat het hier gaat om twee symmetrische cellengroepen, aan de bovenzijde van de Brug gelegen: de Locus Coeruleus (cfr. figuur 8). Zij zenden o.m. impulsen uit die naar de gezichtsbanen leiden, hetgeen waarschijnlijk verklaart waarom visuele beelden in de droom zo'n grote rol spelen; de bewegingsbanen daarentegen worden krachtig geblokkeerd.
Dahlström en Fuxe hebben deze Locus onderzocht onder ultraviolette stralen. Zij vonden er een groene fluorescentie, die de aanwezigheid van nor-adrenaline verraadt. Deze cathecolamine zou dus een rol kunnen spelen in het ontstaan van de droom, zoals de serotonine in het ontstaan van de slaap.
De hypothese van Jouvet kan als volgt geschematiseerd worden (figuur 9): wakker zijn, wanneer de cortex geactiveerd wordt door het reticulair stelsel, inslapen, wanneer de cortex beïnvloed wordt door de serotonine, afgescheiden door de raphé; zij onderbreekt de werking van het reticulair stelsel op de cortex en de gezichtsspieren, niet echter op de tonus van de halsspieren, paradoxale fase, wanneer de nor-adrenaline vrij gemaakt wordt door de Locus Coeruleus; zij supprimeert gedeeltelijk de remmende werking van
| |
| |
het reticulair stelsel op de cortex en de gezichtsspieren,
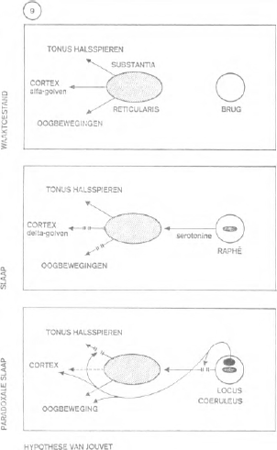
maar blokkeert alle impulsen in de richting van de halsspieren.
Voorlopig is dit de meest gedetailleerde en synthetische theorie over de mechanismen van slaap en droom.
| |
Biologische rol van de droom
Zijn er, tenslotte, fysiologische argumenten die wijzen op het belang van de droom voor ons organisch evenwicht, en misschien zelfs voor ons gedrag? Dement heeft aangetoond dat een proefpersoon die men belet te dromen, veel uitgebreider droomt als men hem daarna ongestoord laat rusten. Het-zelfde fenomeen doet zich voor bij de kat. Men laat een kat slapen op een platformpje midden in een bak water. Zodra een paradoxale fase begint, verliezen de halsspieren hun spanning en de kop komt in contact met het water. De beginnende droom wordt onderbroken, de hals vindt zijn tonus terug. De kop richt zich weer op, tot de volgende paradoxale fase begint. Laat men de kat daarna rustig slapen, dan neemt de paradoxale fase tot 60% van de slaap in, in plaats van de gewone 15 à 20%. De duur van de reconvalescentie hangt trouwens af van de duur van deze tantaluskwelling: een kat die men gedurende 20 dagen belet heeft te dromen, heeft 10 dagen nodig om haar natuurlijk rustritme terug te vinden.
Uit deze twee overeenstemmende observaties concludeert Dement dat de droom wel eens een onmisbare rol zou kunnen spelen in onze fysiologie, al blijft het voorlopig moeilijk zijn juiste functie te definiëren. Recente onderzoekingen van de Joegoslaven Mrsulja, Rakic en Raduvovacki wijzen
| |
| |
in dezelfde richting. Zij constateren dat beroving van paradoxale slaap duidelijk invloed heeft op de hoeveelheid glycogeen (spiersuiker) in de verschillende delen van de hersenen, vooral in de Brug. Wat dit juist betekent, is moeilijk te zeggen, maar de ontdekking is niet te verwaarlozen.
Anderen hebben gewezen op de prikkelbaarheid van individuen die niet kunnen dromen. Misschien mogen we daar echter niet te veel conclusies uit trekken. Wie in om het even welke vastgeroeste gewoonte dag voor dag gestoord wordt, wordt natuurlijk prikkelbaar!
Hoe dan ook, de wetenschap heeft dan toch al een tip gelicht van de sluier die over de geheimen van slaap en droom lag. Misschien wordt het, binnen niet al te lange tijd, wel helemaal duidelijk, waarom de mens, om zichzelf te blijven, slaap en droom niet kan missen.
|
|