|
| |
| |
| |
De bepaling van het geslacht
Door Dr. Theo. J. Stomps.
Een onderwerp, dat in onze eeuw zeer in het bijzonder de aandacht der geleerden tot zich heeft getrokken, is het vraagstuk van de bepaling van het geslacht. Men kan er onder verstaan, eenerzijds het juist begrip daarvan, hoe het optreden van tweeërlei individuen, mannelijke en vrouwelijke, veroorzaakt wordt, anderzijds het onderzoek naar de mogelijkheid, invloed uit te oefenen op de getalverhouding tusschen mannelijke en vrouwelijke individuen. In beide opzichten zijn wij door tal van in de laatste twintig jaar verrichte onderzoekingen een goed eind verder gekomen. Voor verreweg de meeste biologen schijnt het tegenwoordig zelfs eene uitgemaakte zaak te zijn, hoe het komt, dat bij zoovele soorten in de natuur eene scheiding in mannelijke en vrouwelijke individuen plaats vindt. En ten aanzien van de willekeurige bepaling van het geslacht kan niet langer gelden, dat de bij proefnemingen bereikte resultaten ternauwernood de aandacht verdienen.
Zou men eene bespreking over het hier bedoelde onderwerp wenschen te volgen, zoo moet men op de hoogte
| |
| |
zijn van enkele hoofdzaken omtrent den fijneren bouw van planten en dieren. Men moet dan weten, dat deze over het algemeen zijn opgebouwd uit microscopisch kleine hokjes met een wand en een inhoud, waaraan men den naam van cellen heeft gegeven om de gelijkenis van een door een microscoop bezien weefsel met een stuk honigraat. De inhoud der cellen en het voornaamste, wijl eenige levende gedeelte ervan, de z.g. protoplast, omsluit een, bij jeugdige cellen centraal gelegen, lichaam, kern geheeten, dat als het orgaan der erfelijkheid wordt aangemerkt, m.a.w. dat de plaats in de cel zou voorstellen, waar de erfelijke eigenschappen hare stoffelijke basis hebben. Cellen en kernen vermenigvuldigen zich door eene deeling in tweeën en daarbij blijken de kernen weer opgebouwd te zijn uit een, voor elke planten- of diersoort karakteristiek aantal kleinere lichaampjes, de z.g. kernlissen, kernstaafjes, kernsegmenten of chromosomen. Alvorens zich in tweeën te deelen nl. trekt eene kern zich te zamen tot een bepaald aantal compacte lichaampjes, de bedoelde chromosomen, die zich daarop ieder voor zich nauwkeurig in twee gelijke helften splitsen. In de kernen der geslachtscellen, eicellen en spermatozoën, komt, dank zij eene bijzondere kerndeeling, de reduktiedeeling, een aantal chromosomen voor, dat slechts half zoo groot is als dat der lichaamscellen, haploïd is, zooals men het uitdrukt, en bij elke bevruchting wordt het normale of diploïde aantal der chromosomen hersteld, waarin dus elk chromosoom der haploïde serie tweemaal vertegenwoordigd is. Men begrijpt gemakkelijk, dat het wezen der reduktiedeeling daarop berusten moet, dat de twee stellen chromosomen, die bij de bevruchting te zamen kwamen, thans weer gescheiden worden, m.a.w. dat naar de polen der deelingsfiguur uiteen gaan geheele chromosomen, en niet helften van chromosomen, zooals bij de gewone kerndeeling.
Toegerust met deze kennis kan het den lezer niet moeilijk vallen, eene uiteenzetting over kwesties van geslachtsbepaling te begrijpen. Zij moge thans volgen,
| |
| |
gesplitst in twee gedeelten, een eerste, uitvoerig, over de oorzaken van het optreden der geslachten, een tweede, meer beperkt, over de mogelijkheid, op het geslacht invloed uit te oefenen.
Omstreeks het jaar 1891 deed H. Henking de in het eerst ietwat bevreemdende ontdekking, dat de mannelijke individuen van de vuurwants, Pyrthocoris apterus, niet, zooals men zou verwachten, een even, doch een oneven aantal chromosomen in de kernen hebben, dit met het zeer merkwaardige gevolg, dat twee typen van spermatozoën voortgebracht worden. Immers, bij de reduktiedeeling, welke aan de vorming der spermatozoën voorafgaat, zal één der polen van de deelingsfiguur, en daarmede de helft der spermatozoën, bevoorrecht moeten worden, boven de andere, met een extra-chromosoom. Dat Henking juist gezien had, bleek door latere onderzoekingen. In 1899 vond F. Paulmier het oneven chromosoom bij de mannelijke individuen van een ander halfvleugelig insect, Anasa tristis, omstreeks denzelfden tijd Thos H. Montgomery Jr. bij Pentatoma en Protenor, ook al Hemipteren. En niet tot de Hemiptera alleen bleek het verschijnsel beperkt te zijn. W.S. Sutton en C.E. Mc. Clung namen het al spoedig waar bij de rechtvleugelige insecten of Orthoptera, in het bijzonder bij verschillende soorten van sprinkhanen. Van al de genoemde onderzoekers was Mc. Clung de eenige, die de beteekenis van het gevondene eenigermate doorgrondde.
Mc. Clung redeneerde aldus. Blijkbaar hebben de mannetjes een chromosoom te veel. Dit accessorische chromosoom komt slechts in de helft der spermatozoën voor. Dus zullen uit eieren, die door deze spermatozoën bevrucht werden, mannetjes ontstaan. Vrouwelijke individuen treden slechts op als gevolg van eene bevruchting door spermatozoën zonder accessorisch chromosoom. Het extra chromosoom kan gezegd worden door zijne aanwezigheid het mannelijk geslacht teweeg te brengen, het geslacht te bepalen.
Men merke op, dat aan Mc. Clung nog niets met zeker- | |
| |
heid bekend was omtrent de chromosomen van vrouwelijke individuen. Het voortgezet onderzoek leerde nu juist het tegendeel, van wat Mc. Clung gemeend had te mogen aannemen. Het accessorische chromosoom der mannetjes bleek geen extra-chromosoom te zijn.
Aan Miss N.M. Stevens en Edmund B. Wilson komt de eer toe, geheel onafhankelijk van elkander en gelijktijdig in de jaren 1905 en 1906 de waarheid aan het licht te hebben gebracht. Op grond van onderzoekingen bij Hemiptera en kevers kwamen zij tot de conclusie, dat de mannetjes niet een chromosoom te veel hebben, doch een te weinig. Het z.g. accessorische chromosoom is bij de wijfjes (♀) tweemaal, bij de mannetjes (♂) slechts éénmaal vertegenwoordigd. Zoo heeft Anasa t istis ♀ 22 chromosomen, ♂ 21, Protenor belfragei ♀ 14, ♂ 13 chromosomen. De naam ‘accessorisch chromosoom’ moest dus door een beteren vervangen worden, en daarvoor heeft men aanvankelijk gebruikt de termen monosoom, idiochromosoom en heterochromosoom, terwijl men tegenwoordig meestal spreekt van het geslachtschromosoom of x-chromosoom.
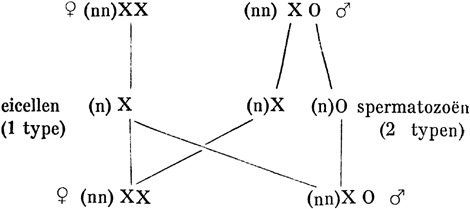
In een eenvoudig schema kan men het verband tusschen de chromosomen en het geslacht tot uitdrukking brengen. Gesteld het haploïde aantal der chromosomen, afgezien van het geslachtschromosoom, zij n. Dan kan men een ♀ individu voorstellen door nnxx en een ♂ door nnxo. De n kan men gemakshalve weglaten, waardoor de formule voor een ♀ individu wordt x x, voor een ♂ x o. De eicellen, met reduktie door een ♀ individu voortgebracht, zal men dan kunnen weergeven door nx, eventueel eene enkele x. Zij zijn alle aan elkander gelijk. De spermatozoën zijn daarentegen van 2 typen, n.l. óf x, óf o. Wordt eene eicel bevrucht door een spermatozoë van het eerste type, dan ontstaat de combinatie x x en dus een ♀, in het tegenovergestelde geval de combinatie x o, die een ♂ zal leveren. Het een en ander voor te stellen aldus:
| |
| |
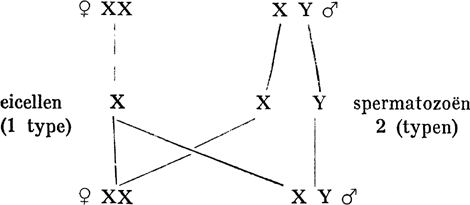
Bij sommige Hemiptera, Lygaeus turcicus b.v. en Euschistus, vond Wilson iets bijzonders, en hetzelfde zag Miss Stevens bij Tenebrio molitor, de meeltor. Ook hier brengen de mannetjes 2 typen van spermatozoën voort en komt in de helft der spermatozoën een x-chromosoom voor. Maar in de andere spermatozoën ontbreekt het niet geheel, m.a.w. vindt men een klein idiochromosoom, dat met het x-chromosoom bij de kerndeelingen in de ♂ individuen een paar vormde. Later is men dit kleine geslachtschromosoom het y-element gaan noemen. Alle eicellen hebben natuurlijk weer het x-chromosoom. Wordt eene eicel bevrucht door een spermatozoë met het x-chromosoom, dan ontstaat een ♀ individu, anders een ♂. Onze schematische voorstelling wordt derhalve:
Het is duidelijk, dat het Protenor-schema en het Lygaeus-schema slechts zeer weinig van elkander verschillen. Laat men het y-chromosoom verdwijnen, dan verkrijgt men terstond den voor Protenor kenmerkenden toestand. Men mag het zóó opvatten, dat het Protenor-type jonger is dan het Lygaeas-type en via dit laatste
| |
| |
ontstaan uit een toestand, waarbij geen verschil tusschen de beide geslachten voorkwam. Inderdaad vond Wilson bij de Hemiptera alle overgangen van soorten, waarbij y ongeveer even groot is als x, naar zulke, waarbij y buitengewoon klein is.
Hiermede hebben wij reeds het voornaamste omtrent de geslachtschromosomen besproken. Verschillende punten vragen evenwel nog onze aandacht, zoo het verder verloop van het onderzoek der geslachtschromosomen, vooral de toestand bij zeeappels, vlinders, amphibieën en vogels, en ten slotte de beteekenis van het onderscheid in de chromosomen tusschen ♂ en ♀ individuen.
Zeer uitvoerig zijn in den loop der jaren de Hemiptera bestudeerd geworden. Wilson wijdde hun eene reeks van wetenschappelijke opstellen, evenzoo Montgomery. Talrijke vormen, in het bijzonder van cikaden, tot de Hemiptera in verband te brengen, onderzocht Alice M. Boring. De waterloopers (Hydrometra) bestudeerde G. Wilke, de ruggezwemmers (Notonecta), die voorzoover onderzocht van het Lygaeus-type bleken te zijn, Ethel Nicholson Browne. Iets wezenlijk nieuws kwam bij al die onderzoekingen ter nauwernood aan het licht. Vermeldenswaardig komt slechts voor de ontdekking, bij verschillende soorten, van samengestelde x-elementen, bestaande uit 2 of meer componenten, die zich intusschen steeds te zamen als eene eenheid gedragen. Het merkwaardigste voorbeeld levert voorzeker Acholla multispinosa, bestudeerd door F. Payne, en waarbij het x-element zich constant voordoet als 5 chromosomen, 2 groote en drie kleine, te zamen nog niet eens zoo groot als het bij deze soort aanwezige y-element. Waar n hier = 10, telt men derhalve in de wijfjes 2 × 10 + 2 × 5 of 30 chromosomen, in de mannetjes, die naast één samengesteld x-element een enkelvoudig y-element hebben, 2 × 10 + 1 × 5 + 1 = 26 chromosomen.
Niet minder veelvuldig dan de Hemiptera werden de Orthoptera op hunne chromosomen onderzocht. In den aanvang trok vooral het werk van Mc. Clung de aan- | |
| |
dacht, voorts een opstel van Montgomery over den sprinkhaan Syrbula. Sprinkhanen werden ook later nog bij herhaling bestudeerd. Regel bleek te zijn, dat de ♂ individuen 1 chromosoom minder hebben dan de ♀. Hetzelfde geldt volgens de onderzoekingen van A. Wassilieff en Max Morse voor de kakkerlakken. Daarentegen bleken ♂ oorwormen aan H. Zweiger, H. Randolph en Miss Stevens naast het x-chromosoom een y-element te bezitten. De krekels eindelijk bestudeerden P. Buchner, S. Gutherz, e.a., niet zonder dat ernstige meeningsverschillen naar voren traden: Buchner meende ook bij de ei-ontwikkeling der wijfjes een accessorisch lichaam te vinden, maar Gutherz wees er op, dat dit lichaam niets met een geslachtschromosoom uitstaande had.
Dat de kevers eensdeels aan het x o-, anderdeels aan het xy-schema beantwoorden, werd boven reeds aangeduid en ook voor de libellen (Odonata) bleek al spoedig hetzelfde te gelden als voor de andere tot dusver genoemde insekten. Anax junius, onderzocht, eerst door Mc. Gill alleen, later door Lefevre en Mc. Gill te zamen, heeft b.v. ♂ 27, ♀ 28 chromosomen.
Eene laatste groep van insecten eindelijk, waarvoor men heeft ingezien, dat aan het chromosomen-complex der ♂ individuen iets ontbreekt, zijn de Diptera, de vliegen en muggen. Het was alweer Miss Stevens, die zich door een onderzoek van verschillende vormen verdienstelijk maakte. Maar hier was het vooral Prof. Th.H. Morgan van de Columbia University te New-York, wiens werk over Drosophila ampelophila (melanogaster) de bananen-vlieg, de belangstelling wekte. Deze vlieg heeft in de lichaamscellen 8 chromosomen, nl. 2 groote haakvormig gebogen paren, 1 zeer klein paar en dan twee geslachtschromosomen, ongeveer 2/3 maal zoo lang als de groote haakvormige chromosomen, en waarvan er één, het y-element, in het ♂ geslacht eveneens haakvormig gebogen is. Zeer talrijke vliegen onderzocht, daartoe gebracht door Morgan's voorbeeld, Ch.W. Metz.
Onze conclusie mag zijn, dat bij de insecten het ver- | |
| |
schijnsel zeer gewoon is, dat de wijfjes één soort van eicellen, de mannetjes twee soorten van spermatozoën voortbrengen, m.a.w. het ♀ geslacht homo-, het ♂ heterogametisch is. Regel is het intusschen geenszins, gelijk wij hieronder nader zullen vernemen. De vraag, of heterogametie van het ♂ geslacht nog elders in het dierenrijk waargenomen is, kan in bevestigenden zin beantwoord worden. De naaste verwanten der insecten onder de gelede dieren, voorts de wormen en de zoogdieren leverden nog vele voorbeelden.
Met de naaste verwanten der insekten worden hier bedoeld eenerzijds de duizendpootachtigen, anderzijds de spinnen. De eerstgenoemden bestudeerden M.W. Blackman en G. Medes, spinnen b.v. Montgomery (Lycosa!), Miss L. Wallace, E.H. Berry.
Aan de wormen werden niet weinige verhandelingen gewijd. Onderzocht zijn in hoofdzaak de Nematoden of draadwormen, waartoe de bekende spoelworm behoort, daarnaast een enkele vertegenwoordiger uit de met de lintwormen verwante groep der Trematoden of zuigwormen. Algemeen bleek het Protenor-schema van Wilson gevolgd te worden. Nadere bijzonderheden gelieve men aan te treffen in een werkje van schrijver dezes over de stoffelijke basis der erfelijkheid bij planten en dieren, dat dezer dagen het licht ziet.
Wat den toestand bij de zoogdieren betreft, zoo heerscht hier over het algemeen veel minder klaarheid dan bij de tot dusver genoemde groepen. Intusschen ziet het er wel naar uit, dat gezegd mag worden, wat voor de Hemiptera enz. gold, nl. dat één type van eicellen en twee typen van spermatozoën worden voortgebracht. Eene waardevolle mededeeling met betrekking tot het Opossum danken wij aan H.E. Jordan, die tevens een overzicht gaf van wat te voren over de geslachtschromosomen der zoogdieren was gewerkt. H. von Winiwarter beschreef heterochromosomen voor de kat, S. Gutherz voor de muis. Zeer veel is er geschreven over de chromosomen van den mensch. W. Flemming bevond in 1898 hun aan- | |
| |
tal te zijn 24. J. Duesberg kwam in 1906 tot dezelfde conclusie, 12 chromosomen tellend in de ♂ geslachtscellen, en dezelfde getallen vond Montgomery in 1912. Hiermede in het geheel niet in overeenstemming zijn de resultaten van von Winiwarter van het jaar 1912. Volgens dezen heeft de vrouw 48 chromosomen, waaronder 2 x-chromosomen, de man 47, waaronder 1 x-element. In de eicellen zouden steeds 24 chromosomen voorkomen, in de spermatozoën deels 23, deels 24. Het zou dus met ons al juist zoo gesteld zijn als met de bovenbedoelde insecten enz., zeer ten gerieve van het ♀ geslacht, dat, gegeven dat de chromosomen algemeen als de dragers der erfelijke eigenschappen gelden, er op zou kunnen bogen, een aantal eigenschappen dubbel te hebben, die bij het ♂ geslacht slechts enkel vertegenwoordigd zijn. De afwijkende resultaten van von Winiwarter hebben intusschen aanleiding gegeven tot het doen van nieuwe onderzoekingen. M.F. Guyer onderzocht negers en blanken en meende in de cellen van de laatstgenoemden een veel grooter aantal chromosomen te vinden, dan in die van negers. Dit klinkt aannemelijk, want neger en blanke staan ver genoeg van elkander af, om een geheel verschillend chromosomenaantal
te hebben. Ook Evans telde bij den blanke 48 chromosomen en wel, in tegenstelling met von Winiwarter, zoowel bij het ♂ als het ♀ geslacht. Men kon dus gaan denken, dat het karakteristieke getal van het blanke ras was 48, al of niet verminderd met één bij het ♂ geslacht, dat voor negers 24. Nieuwe onderzoekingen van Wieman, bij negers en blanken beide, leerden evenwel, dat steeds 24 chromosomen voorkomen. Het ♂ geslacht heeft er evengoed 24, als het ♀, maar één dezer chromosomen is veel te klein, m.a.w. het ♀ geslacht heeft twee x-chromosomen, het ♂ slechts één en daarnaast een y-element, dat zekere eigenschappen mist, die het x-chromosoom wel draagt. De zaak staat dus op het oogenblik zóó, dat het karakteristieke aantal chromosomen van den mensch 24 schijnt te zijn, met deze byzonderheid, dat het ♂ geslacht een
| |
| |
gedeelte van een chromosoom mist, dat bij het ♀ tweemaal vertegenwoordigd is. Von Winiwarter en Evans hebben blijkbaar toevallig materiaal in handen gekregen, dat min of meer abnormaal genoemd kan worden.
Wij zijn thans genaderd tot een nieuw gedeelte van onze bespreking der geslachtschromosomen. Boven werd opgemerkt, dat in het bijzonder de zeeappels, de vlinders, de amphibieën en de vogels ons nog bezig zouden houden. Inderdaad wijken deze met meer of minder groote zekerheid principieel af van de x o- en x y-schemata, die wij tot nu toe steeds van toepassing vonden. Niet het ♂ geslacht is hier, resp. schijnt te zijn, het heterogametische, maar het ♀. Dit laatste brengt dus twee typen van eicellen voort, het ♂ 1 type van spermatozoën.
Dat de zeeappels in dit verband genoemd worden, heeft feitelijk nog slechts historische beteekenis. In 1909 meende F. Baltzer gevonden te hebben, dat voor hen het tegenovergestelde gold als voor Hemiptera, Orthoptera enz. In 1913 trok hij echter zijne bewering terug, nadat inmiddels D.H. Tennent voor den zeeappel Hipponoë heterogametie van het ♂ geslacht had aangetoond.
Van de vlinders evenwel weten wij tegenwoordig met absolute zekerheid, dat de ♂ individuen een zeker geslachtschromosoom twee maal hebben, dat bij ♂ de slechts eenmaal of ten minste niet volledig tweemaal aanwezig is. Reeds lang te voren had men dit uit de resultaten van bastaardeeringsproeven, waarop wij, om niet al te uitvoerig te worden, thans niet willen ingaan, begrepen. Ook hadden verschillende onderzoekers, Miss Stevens, Pauline Dederer, John P. Munson ‘M.H. Cook, Johann Erwin Buder, er op gewezen, dat bij ♂ individuen steeds een even aantal chromosomen voorkomt, zoo bij den wolfsmelkpijlstaart vlgs. Buder 56 in 28 paren. Het bewijs echter werd eerst onlangs geleverd door nauwkeurige onderzoekingen van J. Seiler en L. Doncaster.
De eerste mededeelingen van Seiler hadden betrekking op Phragmatobia fuliginosa, een met den St. Jacobsvlinder en Beervlinder verwante soort van Nachtvlinder.
| |
| |
De mannetjes zijn gekenmerkt door 56 chromosomen, waaronder 2 geslachtschromosomen, de wijfjes hebben er 57, waaronder 1 geslachtschromosoom en eene in twee fragmenten uiteengevallen rest van een 2e geslachtschromosoom, als het ware een samengesteld y-element. Men spreekt intusschen thans niet van een y-element, maar gebruikt in gevallen van heterogametie van het ♀ geslacht voor het geslachtschromosoom en eene eventueele rest daarvan de letters z en w. De mannetjes kan men dus voorstellen door zz, de wijfjes door wz of oz, de spermatozoën, alle aan elkander gelijk en in het bezit van een geslachtschromosoom, door z en de eicellen, van 2 typen, voor de helft met en voor de helft zonder geslachtschromosoom, door w en z resp. o en z. Phragmatobia fuliginosa volgt het wz-schema, alle spermatozoën hebben 28 chromosomen, de eieren voor de helft ook 28, maar overigens 27 + een uit 2 fragmenten bestaand w-element, dus 29 chromosomen.
Vertegenwoordigt Phragmatobia, om zoo te zeggen, het omgekeerde van het Lygaeus-type, den toestand van Protenor, maar dan omgekeerd, heeft men eveneens bij vlinders aangetroffen. Zoo hebben volgens Seiler de wijfjes van Talaeporia tubulosa (eene mot, die een kokertje maakt, vandaar de naam) 59 en de eieren óf 29, óf 30 chromosomen, de mannetjes daarentegen 60 en de spermatozoën steeds 30 chromosomen. En Doncaster, die eene uitvoerige studie maakte van den bessenvlinder, Abraxas grossulariata, vond ten minste bij zekere rassen van deze soort de wijfjes in het bezit van 55, de mannetjes in dat van 56 chromosomen, terwijl bij andere de wijfjes blijkbaar naast het z-chromosoom een w-element hebben.
Behoeven wij, wat de vlinders betreft, niet meer in twijfel te verkeeren omtrent het gedrag der chromosomen, ten opzichte van den toestand bij amphibieën past nog voorzichtigheid in ons oordeel. F. Levy en W.W. Swingle traden op grond van waarnemingen bij kikkers voor de heterogametie van het ♂ geslacht in de bres. R. Goldschmidt kan zich hiermede echter niet ver- | |
| |
eenigen. Hij telde bij een door parthenogenesis, d.i. door kunstmatige ontwikkeling van een onbevrucht ei, verkregen en ♂ uitgevallen kikker 26 chromosomen. In tegenstelling met de verwachting is dit het diploïde aantal. Goldschmidt redeneert nu aldus. De eicel zal, toen de parthenogenetische ontwikkeling haar aanvang nam, het haploïde aantal of 13 chromosomen gehad hebben. Dit aantal werd reeds zeer spoedig, door welke oorzaak dan ook, verdubbeld. Maar dan moeten onder de 26 getelde chromosomen ook twee geslachtschromosomen schuilen en dus moet het ♂ geslacht het homogametische zijn. Voor deze redeneering is zeker veel te zeggen, en het moet worden toegegeven, dat ervaringen, waarover hieronder meer, omtrent de mogelijkheid, bij kikkers en padden invloed uit te oefenen op de getalverhouding tusschen ♂ en ♀ individuen er zeer goed mede in overeenstemming te brengen zijn.
Wat eindelijk de vogels aangaat, zoo kan men er vrij gerust op zijn, dat ook hier het ♀ geslacht het heterogametische is, zulks vooral op grond van de resultaten van kruisingsproeven en niet zoo zeer, omdat wij over veel goed gedocumenteerde gegevens ten opzichte van de chromosomen beschikken. De chromosomen der vogels zijn niet gemakkelijk te bestudeeren, wijl zij eene sterke neiging hebben te zamen te kleven. Intusschen kwamen in 1914 Alice M. Boring en Raymond Pearl met klem op tegen eene vroegere mededeeling van M.F. Guyer, volgens welke de haan en niet de kip twee typen van geslachtscellen zou voortbrengen, en beschreef de laatstgenoemde onderzoeker zelf een tweetal jaren later 2 U-vormige geslachtschromosomen naast 16 gewone voor het ♂ en in het geheel slechts 17 chromosomen, waaronder I U, voor de ♀ individuen van ons gewone hoen.
Na aldus met de voornaamste feiten kennis te hebben gemaakt, keeren wij terug tot de vraag, wat de beteekenis van het onderscheid in de chromosomen tusschen ♂ en ♀ individuen, dat in zoovele gevallen (niet in alle!) werd opgemerkt, zijn kan. En dan kunnen wij vast- | |
| |
stellen, dat de algemeene opvatting tegenwoordig wel schijnt te zijn, dat er eene bepaalde eigenschap is, die beslist, welk geslacht zal optreden, en dat deze eigenschap haar zetel in de geslachtschromosomen heeft. Den gedachtengang van Mc. Clung heeft men voortgezet en met de resultaten van latere onderzoekingen in overeenstemming gebracht. Tot toelichting hiervan diene het volgende, waarbij wij voornl. slechts het oog hebben op organismen als de Hemiptera enz.
Wanneer men te doen heeft met eene soort, waarbij eene scheiding in ♂ en ♀ individuen voorkomt, zoo is het inderdaad niet buitengesloten, dat het van één enkele eigenschap afhangt, welk geslacht te voorschijn zal treden. Mannelijke en vrouwelijke individuen schijnen wel beide de eigenschap voor mannelijkheid te hebben, men denke aan de ♂ trekken, die zoo dikwijls, in het bijzonder bij het ouder worden, bij ♀ individuen zichtbaar worden. Wanneer nu buitendien eene eigenschap voor ♀ worden aanwezig is, krachtig bij de ♀ individuen, zoodat de eigenschap voor mannelijkheid wordt overheerscht, minder krachtig bij de ♂, zoo zoude daarmede het geslacht van deze eene eigenschap afhankelijk kunnen zijn. De vraag is, hoe men zich dat meer of minder krachtig zijn der eigenschap voor vrouwelijkheid moet voorstellen. Daarop kan het antwoord aldus luiden: dat zij in de x-chromosomen zetelt en derhalve bij ♀ individuen tweemaal, bij ♂ slechts eenmaal vertegenwoordigd is.
Deze redeneering, die mutatis mutandis ook op vlinders, vogels enz. van toepassing is, heeft er toe geleid, dat men tegenwoordig meestal als vaststaand aanneemt, dat de geslachtschromosomen het geslacht bepalen. Zoo heel zeker komt dit den schrijver intusschen niet voor. Bewijzen, dat het zoo is, heeft men niet, argumenten voor het tegenovergestelde, n.l. dat het hebben van een of twee geslachtschromosomen een begeleidend kenmerk van het geslacht, een gevolg van het geslacht is, daarentegen te over. Men bedenke b.v. dat veelal geen
| |
| |
onderscheid in de chromosomen tusschen het ♂ en ♀ geslacht bestaat. Wel zou dan de eigenschap voor vrouwelijkheid zeer goed, in plaats van eenmaal afwezig, bij een ♂ individu eenmaal latent of onwerkzaam kunnen zijn, maar hiermede rijst toch de vraag, waarom, bij aanwezigheid van goed gedifferentieerde geslachtschromosomen, de eigenschap voor vrouwelijkheid juist in deze zou moeten zetelen. En wat te denken van gevallen, als dat van de spin Theridium, onderzocht door Montgomery, waarin, krachtens het bovenstaande, het ♂ geslacht t.o.v. de chromosomen heterogametisch is en toch twee typen van eicellen, groote en kleine, worden voortgebracht? Eenige bijzondere gevallen van overerving van het geslacht zijn zeer geschikt, om ons in onze meening te staven.
Ascaris nigrovenosa (Angiostomum nigrovenosum) is een soort van spoelworm, die in twee vormen optreedt, een grooteren, tweeslachtigen, die als parasiet leeft in de long van den kikker, en een kleineren, van gescheiden geslacht, den z.g. Rhabditis-vorm, die met den eerstgenoemden afwisselt en vrij in den bodem voorkomt. Th. Boveri en W. Schleip onderzochten onafhankelijk van elkander, hoe het hier met de chromosomen staat, en kwamen tot overeenstemmende resultaten. De vrijlevende wijfjes en mannetjes hebben resp. 12 en 11 chromosomen, gehoorzamen dus aan het Protenor-schema, de tweeslachtige generatie, die volgt, telt er 12. Eenigermate raadselachtig is het, waarom nu geen individuen met 11 chromosomen optreden. Blijkbaar gaan de helft der spermatozoën der vrijlevende ♂ individuen, waarin het geslachtschromosoom ontbreekt, te gronde. De groote tweeslachtige dieren brengen één type van eicellen met 6 en twee soorten van spermatozoën met 6 en 5 chromosomen voort. Dit moet aldus begrepen worden, dat bij de ontwikkeling der spermatozoën één der beide in de spermatocyt - dat is nog diploïde moedercel der spermatozoën, die de reduktiedeeling doormaakt en daarbij, in den regel, aan vier spermatozoën het aanzijn geeft - nog aanwezige geslachts- | |
| |
chromosomen uitgestooten wordt met het gevolg, dat slechts de helft der spermatozoën een geslachtschromosoom krijgen kan. De redeneering, dat twee geslachtschromosomen een ♀ individu geven, één x-element een ♂ te voorschijn roept, kan hier bezwaarlijk in toepassing gebracht worden, waar individuen met twee geslachtschromosomen ook tweeslachtig kunnen zijn. En het is zeer zeker misplaatst te betoogen, dat bij de vorming der spermatozoën in de tweeslachtige dieren een geslachtschromosoom geëlimineerd wordt en daarom nu
♂ individuen gaan optreden: voor het uitstooten van dat geslachtschromosoom zijn bepaalde eigenschappen noodig en deze zijn het blijkbaar, die het optreden van de ♂ individuen veroorzaken.
Eene zeer merkwaardige overerving van het geslacht vindt men bij bijen en hommels (Apidae), wespen (Vespidae) en mieren (Formicidae), alles vliesvleugelige insecten of Hymenoptera. De algemeene regel is hier, dat een wijfje (koningin) bevruchte en onbevruchte eieren legt, hetzij achtereenvolgens, door het uitgeput raken van den voorraad spermatozoën, die het wijfje bij de paring in een daartoe bestemd reservoir, het receptaculum seminis, opnam, hetzij naar willekeur, door al of niet openen van het receptaculum seminis bij het leggen der eieren. Uit de bevruchte eieren komen dan uitsluitend wijfjes te voorschijn - al naar de wijze, waarop de larven gevoed werden, al of niet (de arbeidsters) van goede geslachtsorganen voorzien - uit de onbevrucht gebleven eieren uitsluitend mannetjes (darren). De eieren zijn steeds van het gereduceerde aantal chromosomen voorzien - bij de honingbij 16 - en dezelfde eieren, die, onbevrucht gebleven, ♂ individuen leverden, zouden na bevruchting wijfjes gegeven hebben. Het ontstaan der mannetjes berust dus op eene echte parthenogenesis, waarbij het haploïde aantal der chromosomen gehandhaafd blijft, en de spermatozoën zullen zonder reduktiedeeling moeten worden voortgebracht. Dat dit inderdaad gebeurt, is door de onderzoekingen van F.
| |
| |
Meves en J. Duesberg, van E.L. Mark en M. Copeland en van H. Nachtsheim bij de honigbij en een paar soorten van wespen, van L. Granata en L. Armbruster bij de bijen Xylocopa violacea en Osmia cornuta, van W. Schleip en H. Lams bij mieren voldoende gebleken. Eene bijzonderheid, overigens van ondergeschikte beteekenis, die daarbij nog voor den dag kwam, is, dat niet 4 goed ontwikkelde spermatozoën uit één spermatocyt ontstaan, zooals de regel is, doch slechts 2 (wespen) of zelfs maar 1 (honigbij).
Niets is nu eenvoudiger, dan aldus te redeneeren: het haploïde aantal chromosomen, waaronder één geslachtschromosoom, geeft een ♂, het diploïde, waaronder twee geslachtschromosomen, een ♀ individu, dus is de beteekenis der geslachtschromosomen als veroorzakers van het geslacht aangetoond. Maar daarbij ziet men over het hoofd, dat het voornaamste punt, dat opgehelderd moet worden, is, dat hier eene geheel normale ontwikkeling steeds weer met het gereduceerde aantal chromosomen doorloopen wordt. Hiervoor zijn stellig bepaalde erfelijke eigenschappen noodig en aanwezig en daartegenover treedt het vraagstuk van de beteekenis der geslachtschromosomen geheel op den achtergrond.
Wij willen van de Hymenoptera geen afscheid nemen zonder met een enkel woord melding gemaakt te hebben van nog merkwaardiger toestanden, gelijk die b.v. bij de Cynipiden of galwespen worden aangetroffen. Van sommige soorten kent men hier slechts wijfjes, die zich langs parthenogenetischen weg voortplanten en zonder reduktiedeeling eieren met het diploïde aantal chromosomen voortbrengen, wat den botanicus aanleiding zou zijn te spreken van eene voortplanting door apogamie (apogamie = afwezigheid van geslachtelijke vermenigvuldiging). Typisch voor Cynipiden is evenwel, dat telkenjare twee generaties met elkander afwisselen, waarvan er een uitsluitend uit parthenogenetisch zich voortplantende wijfjes bestaat en in herfst, winter of voorjaar verschijnt, de andere uit ♂ en ♀ individuen, die in Mei,
| |
| |
Juni of Juli optreden. Nauwkeurig onderzocht is Neuroterus lenticularis en wel door L. Doncaster. De a(po)game generatie van deze soort doet in den herfst aan de onderzijde van eikebladen kleine op knoopjes gelijkende galvormingen ontstaan, die in Maart of April door de zonder uitzondering ♀ veroorzakers worden verlaten. Deze leggen eieren, die zich zonder bevruchting ontwikkelen, in de knoppen van den eik, met het gevolg, dat in Mei aan bladen en katjes ronde sappige galletjes worden opgemerkt (Neuroterus of Spathesaster baccarum), waaruit nu ♂ en ♀ individuen te voorschijn komen. Bevruchting vindt plaats en uit de eieren ontstaat weer de ongeslachtelijke herfstgeneratie. Een agaam wijfje roept óf alleen mannetjes óf alleen wijfjes in het leven. De scherpzinnige lezer begrijpt gemakkelijk, hoe het hier met de chromosomen en de reduktiedeeling gesteld zal zijn. De agame lenticularis-wijfjes hebben het diploïde aantal chromosomen, 20 volgens Doncaster. Een gedeelte brengt zonder reduktie slechts diploïde eieren voort, en daaruit ontstaan de diploïde baccarum-wijfjes, de rest evenwel met reduktiedeeling slechts haploïde eieren, die bij gebrek aan mannetjes niet bevrucht kunnen worden en de haploïde baccarum-mannetjes leveren. De ontwikkeling der spermatozoën gaat, evenmin als bij ♂ bijen en wespen, met reduktiedeeling gepaard, die der door hen te bevruchten eicellen natuurlijk wel.
De galwespen brengen ons op de bladluizen, die eveneens dikwijls galvormingen te weeg brengen en eene merkwaardige levensgeschiedenis hebben. De onderzoekingen van Miss Stevens, W.B.v. Baehr en Th.H. Morgan hebben ons omtrent deze laatste vrij volledig ingelicht. In den nazomer paren ♂ en ♀ individuen met elkander en uit de bevruchte eieren komen in het voorjaar uitsluitend wijfjes te voorschijn. Deze vermenigvuldigen zich langs ongeslachtelijken weg zonder reduktie van het aantal chromosomen een of meer generaties lang, tot weer mannetjes en sexueele wijfjes geboren worden. De vraag is, waarop dit berust en hoe het komt,
| |
| |
dat uit de bevruchte eieren slechts wijfjes verkregen worden.
Wat om te beginnen het tweede gedeelte dezer vraag betreft, het is gebleken, dat bij de bladluizen het ♂ geslacht weer eens gekenmerkt is door het ontbreken van één al of niet samengesteld x-element, dat bij het ♀ tweemaal vertegenwoordigd is. Zoo heeft Aphis saliceti volgens v. Baehr ♀ 6 en ♂ 5 chromosomen, Phyllaphis coweni volgens Morgan dezelfde aantallen, Phylloxera caryaecaulis volgens denzelfden onderzoeker ♀ 8, ♂ 6 en Ph. fallax ♀ 12 en ♂ 10 chromosomen. Men zou dus verwachten, dat steeds twee typen van spermatozoën worden voortgebracht, zulke met en zulke zonder het x-element. In werkelijkheid komen slechts de wijfjes-opleverende spermatozoën tot ontwikkeling en gaan de spermatozoën zonder x-element te gronde. Zeer belangrijk is, dat, wanneer de spermatozoën-moedercel nog ternauwernood met de reduktiedeeling begonnen is, vóórdat de eerste kerndeeling zich voltrokken en het geslachtschromosoom zich hetzij naar de eene, hetzij naar de andere pool begeven heeft, de beide dochtercellen reeds in aanleg zichtbaar zijn, van elkander verschillend in grootte, en het steeds naar de zijde van de grootste cel is, dat het x-element zich begeeft. Dit toch wijst er op, dat niet de spermatozoën wijfjes-gevend werden, omdat het x-element er in terecht kwam, maar dat het geslachtschromosoom er in voorkomt, omdat zij bij de reduktiedeeling tot wijfjes-gevend gestempeld werden. De groote dochtercel van de eerste deeling der reduktiedeeling deelt zich geheel normaal tot twee spermatozoën ieder met een x-element, de kleine degenereert alvorens de tweede deeling voltooid te hebben.
Wij hebben thans begrepen, waarom bij de bladluizen uit de bevruchting slechts ♀ individuen voortkomen, en staan voor de vraag, waarop het berust, dat deze op den duur weer aan mannetjes en sexueele wijfjes het aanzijn geven. Het antwoord is zeer eenvoudig. De sexueele wijfjes ontstaan, evenals tusschengeneraties van onge- | |
| |
slachtelijke wijfjes, zonder dat eenige wijziging in het aantal der chromosomen optreedt, en zijn slechts daardoor gekenmerkt, dat zij hare eieren met reduktiedeeling voortbrengen. De mannetjes evenwel komen voort uit diploïde eieren van ongeslachtelijke wijfjes, waaruit één der beide x-elementen verwijderd werd! En het is alweer eene kwestie van smaak, of men zeggen wil, dat een ♂ individu geboren wordt, omdat één x-element uitgestooten werd, dan wel dat krachtens de erfelijke eigenschappen der soort een mannetje geboren moest worden en daarom een x-element geëlimineerd werd. Wanneer men intusschen met alle verschijnselen rekening houdt, zal men het laatste aan moeten nemen. Bij de genoemde Phylloxera-soorten nl. verschillen de wijfjes-gevende en mannetjes-gevende eieren in grootte, vóór nog van uitstooten van een x-element sprake is; alleen de kleine, blijkbaar dus reeds ♂ eieren stooten een x-element uit en bij Phylloxera caryaecaulis komt er nog deze complicatie bij, dat bepaalde ongeslachtelijke wijfjes slechts groote, andere slechts kleine eieren leggen.
Overzien wij het geheel der hier opgesomde argumenten, zoo ontkomen wij niet aan den indruk, dat het vooralsnog misplaatst zou zijn, de geslachtschromosomen als dragers van het geslacht aan te merken. Men kan werkelijk niet meer zeggen, dan dat zij een begeleidend kenmerk van het geslacht uitmaken. Wat niet wegneemt, dat ons inzicht in het probleem der geslachtsbepaling zich sinds het jaar 1900 ontzaglijk heeft verdiept.
Wij eindigen onze bespreking met eene korte verwijzing naar de resultaten van pogingen, om invloed uit te oefenen op de getalverhouding tusschen ♂ en ♀ individuen, kortweg op het geslacht. Op het eerste gezicht lijkt dit onmogelijk, in het bijzonder voor dengene, die in de geslachtschromosomen de dragers van den, het geslacht bepalenden faktor ziet. Niet waar, de helft der spermatozoën bij de Hemiptera c.s. hebben het geslachtschromosoom, de overige niet, en dus moeten de helft der individuen ♀, de overige ♂ uitvallen? Toch is het in meer dan één
| |
| |
geval reeds mogelijk gebleken, het geslacht te beïnvloeden. Vooral proeven met vlinders en amphibieën gaven overtuigende uitkomsten.
Wat de eerstbedoelde betreft, zoo worde hier nogmaals iets omtrent het werk van J. Seiler medegedeeld. Deze onderzoeker stelde wijfjes van Talaeporia tubulosa, waarvan wij reeds vernamen, dat zij 59 chromosomen hebben, aan abnormaal hooge temperaturen bloot. Het gevolg was, dat veel meer mannetjes uit de door deze ♀ individuen gelegde eieren te voorschijn traden, dan anders het geval zou zijn geweest. Een dergelijk resultaat werd bereikt, door de bevruchting zoo lang, als maar eenigszins mogelijk was, uit te stellen, m.a.w. de eieren overrijp te laten worden, alvorens het toetreden van spermatozoën mogelijk te maken. Willen wij het een en ander trachten te begrijpen, zoo moeten wij ons nog even daarvan rekenschap geven, dat het met de ontwikkeling van eicellen anders gaat dan met die van spermatozoën. Spermatozoën ontstaan, gelijk wij zagen, met zijn vieren uit eene nog diploïde spermatozoënmoedercel. Uit eene zich met reduktie deelende ‘eimoedercel’ of o(v)ocyt ontstaat evenwel slechts een enkele eicel, die de drie andere dochtercellen, welke feitelijk hadden moeten optreden, als z.g. poollichaampjes aan hare oppervlakte uitstoot.
Bij Talaeporia tubulosa hebben de wijfjes 59, de mannetjes 60, de eicellen óf 29 óf 30 en de spermatozoën steeds 30 chromosomen. Uit een ei met 30 chromosomen treedt na bevruchting steeds een ♂ individu te voorschijn. Het is duidelijk, dat een dergelijk ei ontstaat, wanneer het in de ovocyt éénmaal voorhanden geslachtschromosoom voor het ei behouden blijft en niet met de poollichaampjes uitgestooten wordt. Zooveel laat zich dus naar aanleiding van het bovenstaande in elk geval zeggen, dat hooge temperaturen en overrijp worden der eieren - eigenlijk der ovocyten - faktoren zijn, die bevorderlijk zijn voor het binnen het ei blijven van het geslachtschromosoom. Maar of nu het ei met 30 chromosomen tot
| |
| |
een ♂ individu leidt, omdat het geslachtschromosoom er in bleef, dan wel of dit laatste gebeurde, omdat het ei door de genoemde invloeden al tot ♂ gestempeld was, dat zijn vragen, waarop wij, anders dan tegenwoordig te doen gebruikelijk is, het antwoord schuldig willen blijven.
Boven werd er reeds op gezinspeeld, dat het ook bij kikkers en padden mogelijk is gebleken, invloed op het geslacht uit te oefenen, en dit zou steun verleenen aan de opvatting, dat bij deze dieren het ♀ geslacht het heterogametische is. Inderdaad is het aan R. Hertwig en zijne leerlingen G. Kuschakewitsch en E. Witschi, voorts aan Leo Adler gelukt, bij kikkers een doorslaan van de geslachtsverhouding ten gunste der mannetjes en zelfs een algeheel ontbreken van wijfjes te bewerken en wel langs denzelfden weg, dien Seiler later zou betreden, n.l. door de eieren overrijp te laten worden. Wellicht mag daaruit de gevolgtrekking gemaakt worden, dat bij de eiontwikkeling der amphibieën, als bij die der vlinders, een éénmaal aanwezig geslachtschromosoom de keus heeft tusschen in het ei blijven en er met de poollichaampjes uit verwijderd worden, op welke keus de omstandigheden invloed vermogen uit te oefenen. Helen King, die met Bufo lentiginosus, eene Noord-Amerikaansche pad, werkte, bevond de temperatuur en overrijpheid der eieren van geen beteekenis. Daarentegen kon zij, door het water, waarin de eieren afgezet werden, aan te zuren, een overmaat van mannetjes in het leven roepen.
Nog van één verder geval ware gewag te maken, waarin het mogelijk is gebleken, het geslacht te beïnvloeden, terwijl het met zekerheid een organisme betrof, waarbij twee typen van eicellen worden voortgebracht. Dat organisme was de duif en het was O. Riddle, aan wien het gelukte, de normale getalverhouding tusschen ♂ en ♀ individuen te verstoren. De vraag, of ooit bij soorten als de mensch, waarbij de ♂ individuen heterogametisch en, om reden dat zich alle 4 de dochtercellen van een spermatocyt tot levenskrachtige spermatozoën ontwikkelen, van al of niet verwijderen van het x-chromo- | |
| |
soom geen sprake kan zijn, eene verschuiving der geslachtsverhouding bereikt zal worden, moeten wij alweer onbeantwoord laten. Een feit als dat, medegedeeld door Montgomery, nl. dat uit de eieren van de spin Latrodectus 8 maal meer mannetjes dan wijfjes te voorschijn komen, geeft te denken. Maar wellicht heeft de natuur het hier zoo ingericht, dat de spermatozoën zonder x-chromosoom beter voor het bevruchten der eieren geschikt zijn dan de wijfjes-gevende spermatozoën en hebben wij er verder niets achter te zoeken.
Ten slotte de opmerking, dat, waar wij het hier over de mogelijkheid, willekeurig het geslacht te bepalen, hadden, wij slechts het oog hadden op onderzoekingen, waarbij getracht werd, invloed uit te oefenen op de geslachtscellen vóór de bevruchting. Natuurlijk zou men ook kunnen gaan probeeren, de na de bevruchting inzettende ontwikkeling in eene bepaalde richting te leiden. Zulke proeven zijn inderdaad genomen en in sommige gevallen met gewichtige resultaten. Zoo gelukte het aan verschillende onderzoekers, met name aan Witchi, bij kikkers het geslacht te beïnvloeden door na de bevruchting inwerkende faktoren. En onlangs verraste Baltzer de wetenschappelijke wereld met de mededeeling, dat men jonge nog ongedifferentieerde larven van Bonellia viridis, een merkwaardig wormachtig dier uit het zeewater, over welks chromosomen men nog niet ingelicht is, willekeurig kan dwingen tot mannetjes of wijfjes te worden, door hun al of niet de gelegenheid te geven zich vast te zetten op het snuitvormige verlengsel van een volwassen ♀ dier. Met groote belangstelling mag men uitzien naar de resultaten van verdere proefnemingen van gelijke strekking. Het zou uiterst merkwaardig zijn, bijaldien het zou gelukken, een ten opzichte van de chromosomen heterogametisch individu er toe te brengen, onder den invloed van uitwendige omstandigheden, door metagame geslachtsbepaling, zooals men zou kunnen zeggen, het andere geslacht aan te nemen!
|
|
