|
| |
| |
| |
Vijftig jaar kiemplasmatheorie
Een mysterie van onpeilbare diepte ligt omvat in de schakel, die het levend organisme met zijn beide ouders verbindt, een schakel van de minuscule afmeting van een paar duizendste millimeter en van een zeer subtiele structuur, de bevruchte eicel. Ze kwam tot stand door de versmelting van twee voortplantingscellen, bij dieren en mensch eicel en spermatozoïde, bij planten eicel en stuifmeelkorrel, tot één volkomen harmonisch geheel; in die bevruchte eicel ligt alles besloten, wat als erfelijk van de ouders op de kinderen overgaat, de stoffelijke grondslag voor alle erfelijke eigenschappen. En een stap in de ontwikkeling terug gaande, moet deze erfelijke grondslag dus ook in de voortplantingscellen zelf gelegen hebben.
Deze, aan een axioma grenzende, uitspraak werd als onafwijsbaar aanvaard door de Grieksche wijsgeeren en hun geneeskundige landgenooten, en ze geldt nog onverzwakt voor den modernen bioloog. Weliswaar waren onder de Grieken de meeningen verdeeld ten aanzien van de rol, die aan de ‘voortplantingsstoffen’ bij de bevruchting toegewezen is: sommigen van hen geloofden, dat alleen het van mannelijke zijde afkomstige element, het sperma, potentiëel de geheele ontwikkeling van het latere individu bevatten zou, anderen huldigden de opvatting, dat óók van moederszijde iets stoffelijks aan de nakomeling geschonken werd. Wel blijft een dergelijke antithese tot in de achttiende eeuw gehandhaafd, toen ze nog leidde tot de tegenstelling tusschen de ovisten, die het ei als alleen-verantwoordelijk beschouwden, en de animalculisten, die in de spermatozoïden den oorsprong van het nieuwe dier zagen. Wel weten we thans, dat het bevruchtingsproces inderdaad in de versmelting van twee vrijwel gelijkwaardige cellen gekristalliseerd is. Maar de vroegere opvattingen ontkwamen geen van allen aan de noodzakelijke aanvaarding van het axioma der voortplantingscellen als stapelplaatsen van alles, wat erfelijk is, en wij tegenwoordig ontkomen er evenmin aan.
Ondanks haar onaantastbaarheid verbergt deze grondstelling een wereld van vraagstukken, problemen reeds voor de oude Grieken èn problemen voor ons, twintigste-eeuwers. Het meest op den voorgrond treedt hierbij de vraag naar de verhouding tusschen de ouderlijke lichamen en de door de ouders gevormde voortplantingscellen, een vraag, die ook de Grieken als principiëel beschouwden. Indien aangenomen moet worden, dat het ‘sperma’ de overdracht van erfelijke eigenschappen verzorgt,
| |
| |
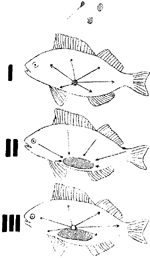
Fig. 1
hoe wordt het dan daartoe in staat gesteld? Twee opvattingen zijn denkbaar, verzinnebeeld in de voorstelling die Johannsen daarvan gegeven heeft (fig. 1): voorop staat in beide de these, dat uit het versmeltingsproduct van eicel en spermatozoïde het geheele lichaam gevormd wordt (I); daarna scheiden de meeningen zich. De eene denkbaarheid laat door alle lichaamsdeelen kleine kiempjes afgeven, die in de geslachtsorganen verzameld worden en door de geslachtsproducten op de nakomeling worden overgebracht om daar weer werkzaam gesteld te worden bij den opbouw van het overeenkomstige lichaamsdeel als dat waarvan ze afkomstig zijn (II). Dan wordt dus een continue reeks van transportprocessen voor deze kiempjes verondersteld: van de ouderlijke lichaamsdeelen naar de geslachtsorganen en van deze weer naar de nieuw-te-vormen lichaamsdeelen van de nakomeling. De andere mogelijkheid is deze, dat de geslachtsstoffen zelf de eigenschappen in één of anderen vorm ‘gepraeformeerd’ bevatten (III) en de voeding deze eigenschappen tot ontwikkeling doet komen. De eerste beschouwingswijze, die we in modernere uitwerking bij Darwin als pangenesisleer terugvinden, werd door de philosophen Leukippos, Empedokles, Demokritos en door de artsen, die aan het Corpus Hippokratikum meewerkten, gehuldigd; de tweede, door Geurts in zijn rijke en rijpe studie over de erfelijkheidsgedachte bij de Grieken (1941) als de ‘trophische’ opvatting gekarakteriseerd, komt misschien bij Anaxagoras en bij Diogenes, maar vooral bij Aristoteles tot uiting.
Twintig eeuwen gaan voorbij, waarin deze problemen vrijwel onherkend blijven sluimeren. De aandacht werd in andere richting geleid: strijd ontbrandde over het axioma zelf. De zeventiende-eeuwsche praeformatisten aanvaardden het in zijn volle consequenties; hun hoofdthema was de vraag, of het mannelijk element, dan wel het vrouwelijke, het eenig noodige voor de ontwikkeling van het jonge individu bevatte: de ovisten, voor wie Harvey en Swammerdam de voornaamste woordvoerders waren, zochten in het ei den aanleg van alle later gevormde organen en eigenschappen, de animalculisten (Leeuwenhoek) meenden de geheele rol van erfelijkheidsoverdrager aan de mannelijke zaad- | |
| |
cel te mogen toekennen. En nadat deze strijd binnen de gelederen der praeformatisten zijn hoogtepunt bereikt had, kwamen als derde partij de verdedigers der epigenesis-theorie (C.F. Wolff) aan de praeformatie-opvatting alle redelijke waarde ontzeggen; voor de epigenetici bestond in den grond eigenlijk geen erfelijkheid, omdat zij meenden, dat alle ontwikkeling van buiten af geïnduceerd werd. Pas nadat in het midden van de negentiende eeuw het bevruchtingsproces als versmelting van de beide ouderlijke elementen, zij het in nog vagen vorm, werd erkend, herkreeg het axioma zijn rechten; de oude Grieksche tegenstelling tusschen pangenisten en trophisten kwam opnieuw naar voren in de beschouwingen, die Herbert Spencer en Charles Darwin eenerzijds, Francis Galton en Hugo de Vries anderzijds aan het erfelijkheidsvraagstuk wijdden.
De fijnzinnige en scherpdenkende wijsgeer Herbert Spencer was de eerste, die in zijn Principles of Biology (1864) uiteenzette, hoe hij zich het wezen der erfelijkheid dacht. Zelf geen bioloog van professie, beschikte hij weinig over eigen ervaring, maar hij verzamelde waarnemingen uit de literatuur om op grond daarvan zijn oordeel over allerlei principiëele vraagstukken der biologie te geven. Daarin legt hij ons een reeks van overwegingen voor over de processen van groei, ontwikkeling, functie, herstel en regeneratie van verloren gegane lichaamsdeelen en hij laat daarop een afzonderlijk aan de verschijnselen en den aard der erfelijkheid gewijd hoofdstuk volgen. Zijn voorafgaande bespiegelingen hadden hem reeds geleid tot het zoeken van een analogie tusschen de eenheden der anorganische wereld, de atomen, en veronderstelbare eenheden van organische natuur, die analoog aan de atomen in anorganische stoffen, als de elementen, waaruit het levende organisme is opgebouwd, door een zekere ‘polariteit’ tot een harmonisch geheel zouden zijn samengeweven. Wat die polariteit precies is, kan hij niet zeggen; hij kan ook geen anderen term ervoor vinden: ‘we may use the term organic polarity or polarity of organic units, to signify the proximate cause of the ability which organisms display of reproducing lost parts... It will be well here to ask what these units are, and by what name they may be most fitly called’ (1864, p. 182).
Als hij nu gaat overdenken, wat voor eenheden het lichaam van het levende wezen samenstellen, dan komt hij allereerst tot de conclusie, dat het geen eenheden van chemische natuur kunnen zijn, want scheikundige stoffen zijn daartoe niet gevariëerd genoeg; de groote veelvormigheid van organismen en hun differentiatie
| |
[pagina t.o. 96]
[p. t.o. 96] | |
Plaat I
Charles Darwin (1809-1884)
August Weismann (1834-1914)
| |
[pagina t.o. 97]
[p. t.o. 97] | |
Plaat II

Hugo de Vries (1848-1935)

Thomas Hunt Morgan (geb. 1866)
| |
| |
in zoo uiteenloopende typen van weefsels, is met behulp daarvan niet te verklaren. Aan den anderen kant lijkt het voor de hand liggend, als de eenheid van het organisme de cel te beschouwen, dus een soort van ‘morphological unit’. Maar ook dit komt hem niet juist voor, want cellen komen, voorzoover hij wist, niet in alle organismen voor, en zelfs als dat het geval zou zijn, dan was deze hypothese nog onaanvaardbaar, ‘since the formation of a cell is, to some extent, a manifestation of this same peculiar power’.
Dus kunnen volgens zijn meening, noch de ‘chemical unit’, noch de ‘morphological unit’ als organische eenheid in aanmerking komen, en dus moet er een tusschending bestaan, dat hij ‘physiological unit’ noemen wil. ‘There seems to be no alternative but to suppose that the chemical units combine into units immensely more complex than themselves, complex as they are; and that in each organism, the physiological units produced by this further compounding of highly compound atoms, have a more or less distinctive character. We must conclude that in each case, some slight difference in their mutual play of forces, produces a difference in the form which the aggregate of them assumes’ (p. 183).
Spencer leidt zoo uit de voorhanden gegevens over groei en regeneratie af, dat het organisme het product is van zulke physiologische eenheden, die aan zijn bouw en zijn differentiatie ten grondslag liggen. En deze eenheden moeten dus ook in de erfelijkheid hun rol spelen; hun lot is door het bovengenoemde axioma bepaald. Spermatozoïden en eicellen zijn dus in wezen niets anders dan voertuigen, waarin groepen van physiologische eenheden van de ouders op de nakomelingen worden overgebracht. Bij de bevruchting worden deze beide groepen van physiologische eenheden, uit spermatozoïde en eicel afkomstig, samengevoegd; in hoofdtrekken zijn deze groepen identiek, maar in ondergeschikte punten verschillen ze van elkaar. ‘They work in unison to produce an organism of the species from which they were derived, but work in antagonism to produce copies of their respective parent-organisms. And hence ultimately results an organism in which traits of the one are mixed with traits of the other’ (p. 254).
Erfelijkheid wordt dus volgens Spencers hypothese door deze physiologische eenheden veroorzaakt, maar bovendien was hij op grond van zijn beschouwingen over aanpassing aan uitwendige omstandigheden overtuigd, dat ook structuurveranderingen, die het gevolg waren van veranderde levensvoorwaarden, op de na- | |
| |
komelingen overgebracht konden worden. Dit vereischte dus nog een hulphypothese, die hij als volgt geeft: ‘Or, bringing the question to its ultimate and simplest form, we may say that as, on the one hand, physiological units will, because of their special peculiarities, build themselves into an organism of special structure; so, on the other hand, if the structure of this organism is modified by modified function, it will impress some corresponding modification on the structure and polarities of its units’ (p. 256).
Spencers opvattingen zijn van zoo groot historisch belang, omdat hij de eerste was, die trachtte voor het geheele erfelijkheidproces een concreten stoffelijken grondslag verantwoordelijk te stellen en zich rekenschap te geven van den aard van dezen grondslag: zijn ‘physiologische eenheden’, die onder gewone omstandigheden het lichaam opbouwen naar het vooraf bepaalde ontwerp, maar onder buitengewone omstandigheden wordt de in het lichaam teweeggebrachte verandering terugwerkend in de physiologische eenheden vastgelegd en zoo op de nakomelingen overgebracht.
Enkele jaren later (1868) volgt Darwin, wiens studie over de ‘Variation of animals and plants’ hem leidde tot een kritische beschouwing van erfelijkheids- en variabiliteitsverschijnselen bij planten en dieren, met het gevolg dat hij een vijftal ‘laws of inheritance’ opstelde: ‘Firstly a tendency in every character, new and old, to be transmitted by seminal and bud generation, though often counteracted by various known and unknown causes. Secondly, reversion or atavism, which depends on transmission and development being distinct powers: it acts in various degrees and manners through both seminal and bud generation. Thirdly, prepotency of transmission, which may be confined to one sex, or be common to both sexes of the prepotent form. Fourthly, transmission, limited by sex, generally to the same sex in which the character first appeared. Fifthly, inheritance at corresponding periods of life with some tendency to the earlier development of the inherited character’ (1868, II, p. 84).
Dat waren voor Darwin vaststaande regels, waaraan de erfelijkheid onderworpen was, en voor die regels trachtte hij een verklaring te vinden door de ‘Provisional hypothesis of pangenesis’, die hij in hetzelfde werk aan zijn mede-onderzoekers ter discussie bood, waarbij uitdrukkelijk het voorloopige krakter van deze hypothese de klemtoon krijgt: ‘It is almost universally admitted that cells, or the units of the body, propagate themselves by self-division or proliferation, retaining the same nature, and
| |
| |
ultimately becoming converted into the various tissues and substances of the body. But besides this means of increase I assume that cells, before their conversion into completely passive or “formed material”, throw off minute granules or atoms, which circulate freely throughout the system, and when supplied with proper nutriment multiply by self-division, subsequently becoming developed into cells like those from which they were derived. These granules for the sake of distinctness may be called gemmules (or pangenes)... Hence, speaking strictly, it is not the reproductive elements, nor the buds, which generate new organisms, but the cells themselves throughout the body’ (1868, p. 374).
Er bestaat dus tusschen de hypothesen van Spencer en van Darwin in zooverre overeenstemming, dat beide het bestaan van stoffelijke eenheden veronderstellen, nl. Spencer physiological units en Darwin gemmules. En bovendien aanvaarden beide de mogelijkheid, dat deze kiempjes in het lichaam veranderd kunnen worden en zoo veranderd in de voortplantingscellen kunnen terug keeren. Beide waren van denzelfden geest doortrokken als de voorstellingen van Leukippos en Hippokrates. Maar Darwins hypothese is meer uitgewerkt en scherper geformuleerd in twee opzichten: ieder celtype vormt volgens hem zijn eigen type van gemmulae, terwijl het begrip van physiologische eenheden door Spencer veel vager gehouden werd; en in de tweede plaats spreekt Spencer zich over het transportmechanisme door het lichaam heen niet uit, waartegenover Darwin het vervoer van zijn gemmulae van de lichaamscellen naar de voortplantingscellen en terug vrij nauwkeurig bepaalt. Dit verschil tusschen beide hypothesen had tengevolge, dat Darwins pangenesis-opvatting wijder verbreid werd niet alleen, maar ook sterker aan critiek blootgesteld werd.
Critiek kwam al spoedig van de zijde van Darwins neef Francis Galton, die zijn aandacht vooral op de erfelijkheid bij den mensch gericht hield. Galton begreep, dat Darwins transporthypothese een toetsing door het experiment mogelijk maakte; indien namelijk deze veronderstelling juist was, dan moesten bij konijnen de vervoerde gemmulae in het bloed aanwezig zijn, en dan moest dus bloedtransfusie van het eene ras in het andere een verandering van hun samenstelling in de voortplantingscellen van dit laatste ras als noodzakelijk gevolg hebben. Deze proefnemingen werden door Galton op groote schaal uitgevoerd (1871); bloed van zilverkonijnen werd in hangoorkonijnen overgebracht en omgekeerd, maar het resultaat was volkomen negatief, de erfelijke aanleg der voortplantingscellen veranderde geenszins, de nako- | |
| |
melingen bleven van het zuivere ouderlijke ras. Daardoor is Galton ertoe gebracht, zelf beschouwingen aan den stoffelijken grondslag der erfelijkheid te gaan wijden; eenige jaren later legt hij ons zijn ‘A theory of heredity’ voor (1875) en nog iets later zijn ‘Typical laws of heredity’ (1877), gevolgd door zijn grootere werk ‘Natural Inheritance’ (1889).
Galton wil gaarne aannemen, dat aan alle erfelijke eigenschappen bepaalde ‘organic units’ ten grondslag liggen, maar hij sluit zich niet aan bij de opvatting van Darwin, dat tusschen deze eenheden geen verband zou bestaan. Ze vormen met hun allen een geheel, dat hij ‘stirp’ noemt. Met het invoeren van deze uitdrukking doet Galton een zeer belangrijken stap terug van de door Darwin aangenomen geheel onafhankelijke kiempjes; in zijn opvatting behooren al deze gemmulae bij elkaar en vormen ze één geheel. Toch neemt ook Galton wel het bestaan van deze eenheden in zijn beschouwingen op: hij aanvaardt ze als grondslag voor groepen van erfelijke eigenschappen, maar hij verwerpt Darwins stelling, dat deze kiempjes door de lichaamscellen gevormd en naar de voortplantingscellen gevoerd zouden worden. In de vier postulaten, die hij als noodzakelijk voor iedere hypothese van organische eenheden beschouwt (1876, p. 331), neemt hij ook deze op, dat de kiempjes zichzelf vermenigvuldigen en bijdragen om de ‘stirp’ van de nakomelingen te vormen. Hij onderscheidt daarbij drie dingen: 1e de ‘stirp’, die een georganiseerde verzameling van een groot aantal kiempjes is, 2e het lichaam met zijn individueele eigenschappen, gevormd uit een klein gedeelte van deze kiempjes en 3e de voortplantingscellen, ontstaan uit de rest van de stirp (1876, p. 343). Er is dus geen sprake meer van een vervoer van gemmulae van de lichaamscellen naar de voortplantingscellen, maar van een doorloopende keten, die de voortplantingscellen van een individu direct verbindt met die van zijn nakomelingen. In Galtons hoofdwerk over erfelijkheid in het algemeen, ‘Natural Inheritance’ drukt hij zich zóó uit: ‘It appears that there is no direct hereditary relation between the personal parents and the personal child, except perhaps through little-known channels of secondary importance, but that the main line of
hereditary connection unites the sets of elements out of which the personal parents had been evolved with the set out of which the personal child was evolved. The main line may be rudely likened to the chain of a necklace, and the personalities to pendants attached to its links’ (1889, p. 19).
Tusschen de beide opvattingen van Darwin en Galton bestond dus een principiëel verschil, dat schematisch door fig. 2 ver- | |
| |
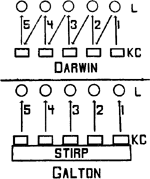
Fig. 2
duidelijkt wordt: uit de ‘kiemcellen’ of ‘voortplantingscellen’ (KC) wordt het lichaam van het organisme (L) gevormd. Volgens Darwin geeft dit weer aanleiding tot nieuwgevormde kiemcellen, waardoor de ‘erfelijkheid van verworven eigenschappen’ wordt mogelijk gemaakt. Volgens Galton danken de kiemcellen, waaruit de individuen der volgende generatie ontstaan, hun oorsprong direct aan het residu van erfelijke substantie; zijn ‘stirp’-begrip zou later in de beschouwingen van August Weismann als ‘kiembaan’ op den voorgrond komen.
De stirp-hypothese bracht voor Galton een moeilijkheid mede, die slechts langs een omweg verklaard kan worden: stamt een organisme van één enkelen ouder af, dan laat zich de stirp als onverbreekbare keten tusschen de voortplantingscellen der opeenvolgende generaties gemakkelijk aannemen; wordt echter een individu uit twee ouders geboren, dan maakt dit een complicatie door een hulphypothese noodig. Om dit te kunnen verklaren, voegt Galton jaren later aan zijn hypothese een andere veronderstelling toe (1897, 1898), volgens welke beide ouders samen de helft bijdragen van het totaal van erfelijke eigenschappen der nakomelingen, de vier grootouders samen een vierde, de acht overgrootouders samen een achtste, enz. Deze wiskundige beredeneering van de samenstelling

Fig. 3
van de stirp werd door Galton (1898) weergegeven in figuur 3, waarin 2 en 3 de aandeelen der ouders, 4, 5, 6 en 7 die der grootouders voorstellen enz. Met deze aanvullende suppositie is Galton al heel weinig gelukkig geweest; ze brengt namelijk als onontkoombare consequentie mee, dat alle kinderen van eenzelfde ouderpaar in stirp, in erfelijken aanleg dus, gelijk zouden zijn èn dat alle door
| |
| |
hetzelfde individu gevormde voortplantingscellen onderling denzelfden erfelijken aanleg zouden overbrengen. Het op Mendels klassieke werk gegrondveste erfelijkheidsonderzoek heeft de onjuistheid van beide consequenties onweerlegbaar aangetoond, en daarmee ook aan deze hulphypothese van Galton haar bestaansrecht ontnomen, waardoor de stirpleer zelf wel niet werd weerlegd, maar toch ernstig verzwakt.
De min of meer speculatieve beschouwingen, die Spencer, Darwin en Galton aan den stoffelijken grondslag van de erfelijkheid wijdden, aanvaarden alle de voortplantingscel als voertuig voor de hypothetische ‘physiological units’, de ‘gemmules’, de ‘stirp’. Maar over de eigenlijke localisatie in de cel, wáár deze erfelijke stof gelegen zou zijn, lieten ze zich niet uit en konden ze zich niet uitlaten, omdat tot 1880 de hiervoor noodige gegevens volkomen ontbraken. Wel was bekend, dat de cel geen homogene eenheid vormt, dat ze integendeel een uitermate gecompliceerde structuur bezit, temidden van een massa eiwitachtig protoplasma een afzonderlijk lichaampje, de celkern, bevat, maar bijzonderheden daarover ontbraken in dien tijd geheel en al; de ware natuur van protoplasma en kern was in ondoorzichtige sluiers gehuld. Een enkele waagt zich met een overmoed, die alleen door een genialen en oorspronkelijken geest gerechtvaardigd kan zijn, tot een uitspraak in deze richting.
Betrekkelijk vaag gehouden is Nägeli's poging (in de jaren tusschen 1865 en 1884), om aan dezen erfelijken grondslag een plaats in de cel aan te wijzen. Zijn uitgangspunt vormt de stelling, dat alle individuen, die tot één soort behooren, gekenmerkt zijn door een aantal gemeenschappelijke kenmerken, waarvoor dan ook in de voortplantingscellen de aanleg aanwezig moet zijn. Die ‘Anlagen’ voor de uitwendig zichtbare eigenschappen liggen volgens zijn opvatting in het protoplasma, doch slechts in een klein deel daarvan. Dit plasma is namelijk geen homogene massa, maar een heterogeen systeem van verschillende bestanddeelen, die door Nägeli met bepaalde namen worden aangeduid: een in water oplosbaar hygroplasma naast een onoplosbaar stereoplasma en dit stereoplasma bestaat dan weer uit doorzichtig hyaloplasma en een ondoorzichtig polioplasma. Dit geheele samenstel van plasmasoorten is doordrongen door een netwerk van ‘idioplasma’, waarin dan de aanleg voor alle uitwendig zichtbare kenmerken gelegen is. Deze fijne strengen of bundels van idioplasma zijn in quantiteit verre de mindere van het overige plasma, waaraan slechts beteekenis voor de voeding toekomt, qualitatief
| |
| |
echter natuurlijk van veel meer beteekenis. Oorspronkelijk alleen in de voortplantingscellen aanwezig, doortrekt dit netwerk langzamerhand het geheele daaruit opgrociende lichaam, zoodat het overal, in elk deel van het individu, zijn werking kan uitoefenen, en ook kan overgaan in de later gevormde nieuwe voortplantingscellen. Ieder individu bezit een in zekere opzichten ‘eigen’ idioplasma; in het algemeen zijn echter de idioplasma-massa's van tot dezelfde soort gerekende individuen onderling zeer sterk gelijkend. Het idioplasma zelf bestaat uit zeer kleine levende deeltjes, ‘micellen’, die een bepaalde rangschikking vertoonen, welke rangschikking ingewikkelder wordt naarmate het organisme ‘hooger’ georganiseerd is. Door dat nu een nieuwgevormde voortplantingscel een hoeveelheid van dit idioplasma meekrijgt en dit idioplasma de ‘Gesamtheit der erblichen Anlagen’ bezit, gaat ook dit aanleg-complex in de voortplantingscellen, dus op de nakomelingen over. Het idioplasma moet dus kunnen groeien, en wel zonder invloed van het daaromheen liggende voedingsplasma, uit zichzelf dus, door intussusceptie. Daarmee vond Nägeli een analogon tot een vroeger door hem onderzocht bestanddeel van de plantencel: de zetmeelkorrel, en daarmee was voor hem het uitgangspunt voor een mechanische voorstelling van de vermenigvuldiging der erfelijke substantie gegeven.
In zijn idioplasma-hypothese tracht Nägeli dus wel een concrete basis binnen de cel te vinden voor de localisatie van de stoffen, die de erfelijkheid veroorzaken, maar hij heeft daarmee geen gelukkige greep gedaan, grootendeels tengevolge van het ontbreken van voldoende feitenmateriaal betreffende den bouw van cel en celkern. Deze fundamenteele gegevens voor verdere gevolgtrekkingen zouden blijven ontbreken tot de jaren tusschen 1873 en 1885, maar desondanks ontmoeten we in de oudere literatuur één uiting, die blijk geeft van prophetischen aard te zullen worden. Het is de geniale intuïtie van Ernst Haeckel geweest, die hem de verschillen in beteekenis tusschen het protoplasma en de celkern deed vermoeden. Hem danken we de volgende uitlating, dateerend van 1866: ‘Wenn wir, wie später gezeigt werden wird, die Form jedes Organismus als das Product aus zwei verschiedenen Faktoren, nämlich aus den ererbten Eigenschaften seiner Materie und aus der Anpassung an die Verhältnisse der Aussenwelt zu betrachten haben, so müssen wir dieses Gesetz auch auf die Elementarorganismen (de kernlooze lagere organismen, zooals bacteriën) anwenden können. Hier scheinen nun die beiden Funktionen der Erblichkeit und der Anpassung bei den kernlosen Cytoden noch nicht auf diffe- | |
| |
rente Substanzen vertheilt zu sein, sondern der gesammten homogenen Materie des Plasma zu inhärieren, während dieselben bei den kernführenden Zellen (der hoogere organismen) in der Weise auf die beiden heterogenen activen Substanzen der Zelle vertheilt sind, dass der innere Kern die Vererbung der erblichen Charaktere, das äussere Plasma dagegen die Anpassung, die Accomodation oder Adaptation an die Verhältnisse der Aussenwelt zu besorgen hat ... Wenn wir demgemäss das Plasma vorzugsweise als den nutritiven, den Nucleus (celkern) dagegen vorzugsweise als den reproductiven Bestandtheil
der Zelle ansehen können, und wenn wir dazu den im fünften Buch nachgewiesenen Zusammenhang einerseits zwischen der Ernährung und Anpassung, andererseits zwischen der Fortpflanzung und Erblichkeit in Erwägung ziehen, so werden wir mit Recht den Kern der Zellen als das hauptsächliche Organ der Vererbung, das Plasma als das hauptsächliche Organ der Anpassung betrachten können’ (1866, I, p. 287-288).
Deze heldere en duidelijk geformuleerde uitspraak steekt wel bijzonder scherp af bij alle in dien tijd geldende min of meer vaag gehouden erfelijkheidstheorieën en is zeker wel een uitnemend voorbeeld van wat een vooruitziende geest als die van Haeckel zonder heel veel feitenmateriaal als een juiste en gerechtvaardigde hypothese kan aanvoelen.
Dit feitenmateriaal werd in de jaren 1873 en volgende bijeengebracht en het zijn vooral drie momenten, die hierbij een rol gespeeld hebben.
Daarbij staat voorop de ontdekking in 1875 van het verloop van het bevruchtingsproces, het versmelten der beide voortplantingscellen, van zoölogische zijde door Oscar Hertwig, van botanischen kant door E. Strasburger. Dit leidde tot de conclusie, dat het essentiëele van de bevruchting gelegen is in een versmelting van de beide kernen der voortplantingscellen, terwijl aan het protoplasma, dat immers in de mannelijke spermatozoïde niet of in zeer kleine hoeveelheid aanwezig is, geen beteekenis bij dit proces kan worden toegekend. Het vermoeden bleek dus gewettigd, dat de kern het monopolie zou bezitten ten opzichte van het overbrengen der erfelijke eigenschappen, of althans een hoofdrol daarbij zou spelen. Dit bleef echter nog lang een, weliswaar stevig gegrond, vermoeden. Eerst in 1909 werd het exacte bewijs voor deze stelling geleverd door C. Herbst, die erin slaagde de celkern van een onbevrucht zeeëgelei zich in tweeën te laten deelen en één dezer beide eikernen door één spermatozoïde te laten bevruchten, terwijl de andere onbevrucht
| |
| |
bleef. Het resultaat was ondubbelzinnig: uit deze eicel groeide een larve op, die uit twee helften bestond; de ééne helft toonde eigenschappen van beide ouders, de andere helft alleen eigenschappen van de moeder afkomstig. Daarmee was dus wel onweerlegbaar aangetoond, dat de kern van de spermatozoïde erfelijke eigenschappen had overgebracht, waardoor dus Haeckels veronderstelling van 1866 volkomen bevestigd werd.
Van vèrstrekkende beteekenis bleek ook het onderzoek naar den bouw van de celkern zelf; evenals de cel, bleek ook de celkern een zeer bepaalde structuur te bezitten. Ze maakt periodiek belangrijke veranderingen door: afwisselend een rusttoestand en een deelingsstadium. De term ‘rusttoestand’ is eigenlijk in hooge mate misleidend: er hebben inderdaad in dit tijdsverloop zeer belangrijke levensprocessen plaats, waarin een innig contact tusschen den inhoud van de celkern en het omgevende protoplasma wordt tot stand gebracht, maar de term is eenmaal ingeburgerd als tegenhanger van de kerndeelingsprocessen. En juist door het onderzoek van die deelingsprocessen is de bouw van de kern ons duidelijk geworden; immers tijdens de voorbereiding voor zulk een kerndeeling kunnen door bepaalde kleurmiddelén allerlei inwendige details van den kernbouw zichtbaar gemaakt worden, die gedurende den rusttoestand aan onze waarneming ontsnappen. Van het grootste belang blijkt dan het feit, dat in de celkern een aantal paren van uiterst kleine draadjes, van eenige duizendsten millimeters lengte, aanwezig zijn. Het aantal paren van zulke draadjes, ‘kernlissen’ of ‘chromosomen’ genoemd, is constant en voor de soort, waartoe het individu behoort, typeerend: de mensch bezit in al zijn lichaamscellen 24 paar chromosomen. Twee verschijnselen trokken nu al spoedig de aandacht van hen, die zich in die jaren omstreeks 1880 met deze onderzoekingen bezig hielden, verschijnselen, die als tweede en derde moment hun rol zouden spelen in de erkenning van de stoffelijke basis der erfelijkheid.
In de eerste plaats het feit, dat deze draadvormige chromosomen vóór de kerndeeling met buitengewoon groote zorgvuldigheid in twee overlangsche helften gespleten worden, zoodat na de kerndeeling van een lichaamscel ieder der beide nieuwgevormde kernen weer precies, en zéér precies, hetzelfde stel chromosomen heeft gekregen, als de oorspronkelijke celkern bevatte; de stoffen, die deze chromosomen opbouwen, bleken in twee volkomen gelijke hoeveelheden verdeeld te zijn.
En het tweede punt, dat de aandacht trok, was dit, dat de vorming van de voortplantingscellen uit een lichaamscel iets bijzon- | |
| |
ders vertoont: de beide chromosomen, die een paar vormen, wijken bij deze kerndeeling uiteen, waarbij de eene in de eene voortplantingscel terecht komt, de tweede in de andere, zoodat bij den mensch elke voortplantingscel niet 24 paar, maar 24 enkele chromosomen meekrijgt. Deze zoogenaamde reductiedeeling, waardoor het aantal chromosomen in de kiemcellen tot de helft wordt gereduceerd, komt zoowel voor bij het ontstaan van eicellen, als bij de vorming van spermatozoïden. Versmelten nu twee voortplantingscellen met elkaar (een eicel en een spermatozoïde) en dus ook hun beide kernen, dan is het resultaat, dat weer een celkern gevormd wordt met de chromosomen in paren, waardoor het voor alle lichaamscellen kenmerkende chromosomenaantal hersteld is.
Deze drie gegevens waren voor de daarbij betrokken onderzoekers (E. van Beneden, Th. Boveri, W. Flemming, O. en R. Hertwig, A. Koelliker, C. Rabl, W. Roux, E. Strasburger, W. Waldeyer) aanleiding om vooral aan de stof, waaruit die chromosomen zijn gebouwd, de ‘chromatine’, waarde te hechten als de eigenlijke materiëele grondslag der erfelijkheid. Koelliker bijv. zeide in 1885 (p. 21): ‘Das Idioplasma der zeugenden Kerne ist offenbar an eine chemisch und morphologisch typische Substanz - nennen wir dieselbe der Kürze halber Chromatin oder Nuklein - gebunden, die nie im Zelleninhalte, wohl aber in allen Kernen ohne Ausnahme gefunden wird’. En Strasburger drukt het nog scherper uit: ‘Die Zusammensetzung des Kernfadens aus Stücken, welche verschiedenen Generationen des betreffenden Organismus entstammen, machen ihn zum Wächter über die specifischen Eigenschaften des Organismus besonders geeignet’ (1884, p. 145).
Zoo nadert het jaar 1892, waarin August Weismann zijn reeds vroeger gedeeltelijk en in voorloopigen vorm gepubliceerde beschouwingen over de erfelijkheidsvraagstukken zou samenvatten in zijn monumentale werk ‘Das Keimplasma’.
Weismann was als het ware aangewezen om in dit zoo moeilijk doorzichtige problemencomplex eenig licht te brengen. Hij beschikte over een rijke, door eigen onderzoek verkregen, ervaring op biologisch gebied. Zijn wetenschappelijk werk begon met enkele kleinere physiologisch-chemische publicaties, gevolgd door uitvoeriger studies over histologische onderwerpen. Daardoor kwam hij van de geneeskunde naar de zoölogie, om zijn verdere leven geheel hieraan te wijden. Hier betrad hij het klassieke, door Malpighi en Swammerdam geopende terrein: de pro- | |
| |
cessen van de embryologische ontwikkeling en van de metamorphose der levensphasen van insecten boeiden hem zoozeer, dat hij geen geestelijke of lichamelijke inspanning vermeed, zijn oogen veel te weinig spaarde. Met het noodlottig gevolg, dat hij op dertigjarigen leeftijd zijn microscopische werk onderbreken moest; eerst tien jaar later herstelde zijn gezichtsvermogen zich voldoende; tien jaar rust hergaf hem de mogelijkheid zijn eigenlijke werk te hervatten. Een tweede, eerbiedwekkend actieve periode van tien jaar (1874-1884) volgde, waarin hij vooral de levenswijze, voortplanting en ontwikkeling van waterbewonende dieren, watervlooien en kwallen, als onderwerpen voor zijn waarnemingen koos. Maar na tien jaar van rusteloozen arbeid moest hij zich opnieuw aan het noodlot onderwerpen; zijn oogen werden zooveel slechter, dat eigen microscopische waarneming een onmogelijkheid werd.
Intusschen was de eerste periode van gedwongen rust evenmin als de rest van zijn leven, een tijd van onwerkzaamheid; juist door het gedwongen afstand doen van microscopisch onderzoek werd zijn geestelijke activiteit in andere richting geleid: de brandende problemen van de afstammingsleer, door Darwins werk (1859) zoo plotseling in het centrum geplaatst, grepen ook Weismanns geheele persoonlijkheid; zijn waarnemingsvermogen richtte zich op andere objecten, zijn geest op evolutievraagstukken. Als hij in 1868 zijn ambt van hoogleeraar aan de universiteit te Freiburg aanvaardt, houdt hij een rede over ‘Die Berechtigung der Darwinschen Theorie’; daarin ligt het uitgangspunt voor een reeks van ‘Studien zur Deszendenztheorie’. Zijn analyse van het complex van problemen, dat de evolutie ons voorlegt, leidde tot een scheiding tusschen de processen van variabiliteit, van erfelijkheid, van selectie en van correlatie, en daaronder ziet hij het erfelijkheidsvraagstuk weer als een weefsel, waarin de verschijnselen van het ontstaan en het rijpen van voortplantingscellen en van de voortplanting in al haar verschillende uitingen het stramien vormt, dat als grondslag voor de daardoorheen gevlochten erfelijke eigenschappen dient. Vijftien jaar later (1883) begint hij zijn prorectoraatsrede te Freiburg met de indrukwekkende woorden: ‘Von der Vererbung möchte ich reden, diesem Grundpfeiler alles Beharrungsvermögens der organischen Formen, dem unbefangenen Laien so selbstverständlich und keiner besonderen Erklärung bedürftig, der Reflexion so verwirrend durch die unendliche Mannigfaltigkeit ihrer Aeusserungen, und so rätselvoll ihrem eigentlichen Wesen nach’.
Twee uitgangspunten vindt Weismann voor zijn beschou- | |
| |
wingen: het probleem der sterfelijkheid van levende wezens èn de ontwikkelingsprocessen, waardoor de voortplantingscellen gevormd worden.
Ieder individu van een soort van hoogere planten of dieren heeft een voor die soort typeerenden, beperkten levensduur, moge deze een deel van een dag of eenige eeuwen omvatten. Maar in dit opzicht bestaat er een scherpe tegenstelling tusschen deze ‘hoogere’ organismen, wier lichaam uit een kleiner of grooter aantal cellen is opgebouwd, en de ‘lagere’ eencelligen, waarvan het ‘lichaam’ uit één enkele cel bestaat. Een eencellige deelt zich; twee organismen ontstaan en vermenigvuldigen zich op hun beurt weer, en nogmaals, en nogmaals, totdat uit de ééne enkele cel een ontelbaar aantal cellen, die elk een volkomen zelfstandig onafhankelijk individu vormen, geboren is. Het oorspronkelijke individu, de oorspronkelijke cel is verdwenen, maar niet gestorven; een ééncellige kàn door den dood vernietigd worden, maar ze is, indien geen fatale omstandigheden ingrijpen en de levensvoorwaarden gunstig blijven, potentiëel onsterfelijk. De cel zelf fungeert tegelijkertijd als lichaam èn als voortplantingscel; een scheiding tusschen ‘lichaam’ of ‘soma’ en ‘kiemcel’ of ‘germa’ wordt bij de eencelligen niet aangetroffen.
Bij de meercellige organismen komt echter zulk een scheiding van lichaamscellen en voortplantingscellen op den voorgrond: de lichaamscellen differentiëeren meer en meer in verband met de hun opgedragen functie, hun werkzaamheid wordt een physiologische, hun potenties beperkt. Het reproductievermogen ten dienste van het organisme wordt aan een bijzondere groep van cellen toegewezen, aan de kiemcellen, die met de speciale taak van de voortplanting belast zijn.
Dit principiëele verschil tusschen soma en germa, dat bij alle meercellige planten en dieren voorhanden schijnt te zijn, vindt zijn uitdrukking in de wijze, waarop de voortplantingscellen gevormd worden. En daarmee krijgt Weismann zijn tweede gegeven ter beschikking. Zijn eigen uitvoerige en nauwkeurige studies over het ontstaan van de voortplantingscellen bij kwallen hadden hem geleid tot de theoretische gevolgtrekking, dat alleen zeer bepaalde eenheden onder de groote massa van cellen, die uit de bevruchte eicel door herhaaldelijk opeenvolgende deelingen ontstaan, in staat zijn voortplantingscellen te vormen. Deze speciale cellen, de toekomstige producenten van kiemcellen dus, komen in de ontwikkelingsperiode tusschen bevruchte eicel en volwassen dier reeds op een vrij vroeg stadium afzonderlijk te liggen; aan den verderen opbouw van het lichaam nemen zij geen deel; ze worden
| |
| |
als het ware in reserve gehouden. Weismann grondt hierop de conclusie, dat de stoffelijke grondslag van de erfelijke eigenschappen, die hij ‘kiemplasma’ noemt, in volledigen staat alleen aan deze toekomstige kiemcellen wordt meegegeven, terwijl de overige voor lichaamscellen bestemde gedeelten van het ontwikkelende dierenlichaam slechts gedeelten van dit kiemplasma ontvangen.
Door deze overwegingen bereikt Weismann de opvatting, die ook reeds door Galton in zijn stirptheorie was vastgelegd: het ‘kiemplasma’ vormt een doorloopende keten, die generatie na generatie met elkaar verbindt; het kiemplasma wordt van ouders op nakomelingen in onveranderden staat overgedragen en reeds spoedig na de eerste ontwikkeling van de bevruchte eicel als reservevoorraad voor de later te vormen voortplantingscellen op een zijspoor gerangeerd.
Zoo vindt Weismann ook een aanknooping tusschen zijn beide uitgangspunten. Bij de ééncelligen is lichaamsplasma en kiemplasma in dezelfde cel, die alleen het individu vormt, vereenigd; de eencellige is onsterfelijk. In het lichaam van de meercelligen is tusschen lichaamsplasma in de cellen, die voor lichamelijke functies dienen, en kiemplasma, dat in de toekomstige voortplantingscellen is ‘opgeslagen’, een scherpe scheiding tot stand gekomen; de lichaamscellen zijn sterfelijk, aan het kiemplasma kan een onsterfelijke natuur worden toegekend.
Tot op zekere hoogte loopen dus Galtons stirptheorie en Weismans kiemplasmatheorie aan elkaar evenwijdig; maar van dit punt af wijken hun opvattingen uiteen. Galton had een speciale constructie voor de stirp voorgesteld, die opgebouwd zou zijn uit de stirps van alle voorouders. Weismann was door de ter beschikking staande gegevens van het celkernonderzoek in staat, zijn theoretische beschouwingen verder uit te breiden, beter te fundeeren, en de voor Galton onoverkomelijke moeilijkheden te overwinnen. Hij was bekend met de rol, die de chromosomen bij de kerndeeling spelen, hij begreep de beteekenis van de nauwkeurige overlangsche splijting der chromosomen tijdens de kerndeeling, en van de halveering, waaraan hun aantal gedurende de reductiedeeling, die tot de vorming van voortplantingscellen leidt, onderworpen wordt. Hij maakte deel uit van de groote groep van grootendeels Duitsche biologen, die in de jaren tusschen 1870 en 1890 dit belangwekkende terrein van celonderzoek ontgonnen hadden, en hij zocht voor dit biologische probleem een biologische verklaring.
In de lijn, waarin Nägeli een bepaald deel van de cel als de
| |
| |
stoffelijke basis voor de erfelijkheid meende te kunnen veronderstellen, zijn idioplasma, dat het geheele protoplasma zou doortrekken, zoekt ook Weismann een concreten grondslag voor het kiemplasma. Gelijktijdig met de uiteenzettingen van Oscar Hertwig, van Koelliker en van Strasburger, geeft hij in 1885 een eerste ontwerp van zijn kiemplasmahypothese: ‘Die Kontinuität des Keimplasmas als Grundlage einer Theorie der Vererbung’. Maar in tegenstelling tot Nägeli en als nadere uitwerking van de gedachten der anderen, localiseert Weismann dit kiemplasma met groote scherpte in de chromosomen. Alle levende stof, dus ook het plasma van cel en kern, denkt hij zich samengesteld uit zeer kleine eenheden, levensdragers, biophoren, die hij als groepen van moleculen beschouwt, en die door het absolute getal moleculen, dat ze bevatten, door de verhoudingsgetallen der verschillende moleculen, hun chemische samenstelling en eventueele isomerie en hun groepeering uitermate uiteenloopend in karakter kunnen zijn. Het zijn volstrekt geen hypothetische eenheden, meent hij; ze moeten bestaan, want de verschillende levensverschijnselen moeten aan verschillende vormen van substraateenheid gebonden zijn. Deze biophoren zijn overal in het levende organisme aanwezig, zoowel in het als stofwisselingsterrein werkende overige protoplasma, als in het kiemplasma der chromosomen. In aansluiting aan een beschouwing van Hugo de Vries (1889) over Intracellulaire pangenesis neemt Weismann aan, dat deze biophoren uit de celkern naar buiten kunnen gaan, en in het de kern omgevende protoplasma hun rol kunnen spelen. Biophoren veroorzaken dan in het celplasma een specifieke werking, hetzij een physiologisch proces of een vormende functie. Voor iedere cel of groep van gelijke cellen, die gedifferentiëerd wordt, is een speciale groep van biophoren aansprakelijk, die met elkaar de ontwikkeling van dit celtype (spiercellen, hersencellen, beendercellen,
kliercellen, enz.) determineeren; de groep van biophoren wordt dan als ‘determinante’ gekenmerkt. Deze determinanten kunnen één of meermalen in het kiemplasma aanwezig zijn; ze zijn daarin op vaste plaatsen gelocaliseerd en groepsgewijs opgesloten in ‘iden’, schijfvormige lichaampjes, reeds door anderen in de chromosomen waargenomen en als ‘chromatine-corpuscula’ beschreven. Zulk een ide, die het totale determinantencomplex zou bevatten, vormt dus den erfelijken grondslag van het geheele individu in vastomschreven rangschikking. Elke ide is van een afzonderlijken voorouder afkomstig, als zoogenaamd ‘Ahnenplasma’, en het verschil tusschen de afzonderlijke iden veroorzaakt dus de individueele verschillen, die tusschen de
| |
| |
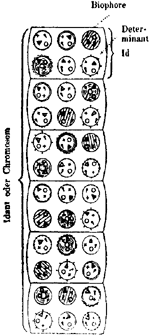
Fig. 4
voorouders bestonden. Een aantal schijfvormige iden is aan elkaar verbonden tot een chromosoom, door Weismann ‘idant’ genoemd. Schematisch wordt Weismanns voorstelling van den bouw van het chromosoom als grondslag der erfelijke substantie in figuur 4 weergegeven.
Een dergelijke constructie van het chromosoom als complex van ‘erfelijkheidsdragers’ legt ons twee vragen voor: is de gewone kerndeeling, waarbij de chromosomen zorgvuldig in twee helften overlangs gespleten worden, in staat de gelijkheid in erfelijken aanleg tusschen de beide ‘dochterkernen’ te verklaren? En: hoe kan het opeenhoopen van steeds meer ‘Ahnenplasma’-massas, die de Ide vormen, vermeden worden? De eerste vraag blijft tot in onzen tijd een van de moeilijkste problemen, moeilijk voor Weismann, moeilijk ook voor ons, die over eenig inzicht in den physisch-chemischen bouw van het chromosoom denken te beschikken. Maar Weismann bezag dit probleem van een anderen kant: hij trachtte het in verbinding te brengen met het zoo belangrijke en nog steeds zoo veelomstreden vraagstuk en de differentiatie, waardoor verschillende typen van organen van weefsels uit theoretisch in aanleg identieke cellen gevormd worden. Voor de oplossing van dit probleem vond Weismann een concreet gegeven. Onderzoekingen van van Beneden over de ontwikkeling van het bevruchte ei van een spoelworm hadden aangetoond, dat hier van de eerste celdeelingsstadia af een ‘chromatinediminutie’ plaats heeft; hij kon bewijzen, dat er reeds dadelijk bepaalde cellen opzij gezet worden, waaruit de latere voortplantingscellen ontstaan en waarin het chromosomenbezit volledig blijft, terwijl de andere cellen altijd een chromatinevermindering ondergaan, zoodat de lichaamscellen een onvolledig bezit van het chromosomencomplex erven. Dit proces leidt tot de mogelijkheid van het opstellen van een ‘kiembaan’ als bron voor de toekomstige voortplantingscellen eenerzijds en daarnaast van celreeksen voor bepaalde embryonale regionen en de daaruit opgroeiende organen anderzijds. Tijdens de lichamelijke ontwikkeling zouden dus voortdurend determinanten afgesplitst worden, waardoor tenslotte elk orgaan een bepaalde determinantengroep en elk
orgaandeel bepaalde determinanten toegewezen krijgt. Deze uitwerking van Weismanns opvattingen, de
| |
| |
hypothese der zoogenaamde ‘erbungleiche Teilung’ wordt tegenwoordig weinig meer aanvaard; of ze inderdaad zoo onaannemelijk is, als thans gemeend wordt, zal de toekomst moeten uitwijzen.
Het tweede punt, de ophooping van massa's van Ahnenplasma, het punt, waarop Galton tenslotte schipbreuk heeft geleden, kon door Weismann met grooter zekerheid worden beantwoord: de reductiedeeling halveert het aantal chromosomen, halveert dus ook het aantal iden en voorkomt een te groote agglomeratie van voorafgaande Ahnenplasmas. In dit opzicht is Weismanns interpretatie nog heden aanvaardbaar.
Zoo heeft Weismann ons in 1892 de meest uitgewerkte hypothese geschonken betreffende de localisatie van den erfelijken grondslag in de cel, in de celkern, in de chromosomen; zijn hypothesencomplex geeft een voor dien tijd volkomen aannemelijk beeld van de structuur der chromosomen als stapelplaatsen, waar de materiëele grondslag der erfelijke eigenschappen ligt bewaard.
We leven thans vijftig jaar later. Vijftig jaar, waarin veel feiten zijn toegevoegd, veel hypothesen door andere zijn vervangen, veel verhelderd, veel onopgelost gebleven. Het onderzoek is verder gegaan: het probleem van de stoffelijke basis der erfelijkheid door experimenten aangevat, door steeds meer verfijnde microscopische techniek in bewerking genomen.
De hypothetische eenheden, Spencers physiological units, Darwins gemmules, Weismanns biophoren, zijn thans vervangen door ‘genen’, die elk voor zich niet op een orgaan, een celgroep of een cel betrekking hebben, maar op een duidelijk-omschreven eigenschap van het organisme. Reeds in 1889 heeft Hugo de Vries de pangenesis-hypothese van Darwin aan een ontleding onderworpen; hij scheidde met groote scherpte de veronderstelling der erfelijke eenheden van hun transportmogelijkheid. ‘I. In den Zellen gibt es zahllose, unter sich verschiedene Theilchen, welche die einzelnen Zellen, Organe, Funktionen und Eigenschaften des ganzen Individuums vergegenwärtigen. II. Ausserdem sondern aber die Zellen des Organismus in jedem Entwicklungsstadium solche Theilchen ab, welche den Keimzellen zugeführt werden, und diesem jene Eigenschaften mitteilen, welche die betreffenden Zellen während ihrer Entwicklung etwa erworben haben’ (1889, p. 62-63). Het tweede gedeelte verwerpt de Vries, zooals velen en vooral ook Galton en Weismann vóór hem gedaan hadden, volkomen; in het eerste ge- | |
| |
deelte verwijdert hij de afzonderlijke cellen, organen en functies en beperkt het begrip van erfelijke eenheid tot den stoffelijken grondslag van een eigenschap. Het is de vraag, of de Vries hier Darwins hypothese niet tegen Darwins eigen opvatting in heeft veranderd. Immers deze laatste erkent in 1888 (Animals and Plants, sec. ed., p. 350): ‘This hypothesis has been severely criticised by many writers’ en in antwoord aan Galton, wiens experimenten negatief waren uitgevallen: ‘I certainly should have expected that gemmules would have been present in the blood, but this is no necessary part of the hypothesis, which manifestly applies to plants and the lowest animals’; hij handhaaft echter zijn uiteenzettingen van 1868 in onveranderden vorm. Maar hoe dit ook zij, de Vries heeft zeker door zijn eigen
interpretatie van Darwins hypothese den inhoud daarvan van ernstige gebreken bevrijd en de kern zoo samengevat, dat ze nog heden aanvaard kan worden. ‘Es sei mir gestattet, die hypothetischen Träger der einzelnen erblichen Anlagen mit einem neuen Namen zu belegen und Pangene zu nennen’.
Dat inderdaad het bestaan van pangenen, of genen, zooals ze thans genoemd worden, voor elke erfelijke eigenschap, aanvaardbaar is en zelfs wel als bewezen beschouwd mag worden, danken we aan het werk van G.J. Mendel, die in 1865 zijn baanbrekende, maar lang onbekend gebleven publicatie over kruisingsproeven met erwten in het licht gaf. Zijn oorspronkelijke geest, zijn heldere formuleering van een enkele te beantwoorden vraag, zijn inzicht in de alleen-mogelijkheid van een oplossing door een doelbewuste proefneming, zijn scherp-wiskundige behandeling en analyse der verkregen gegevens en zijn logische interpretatie hebben inderdaad de conclusie ten volle gewettigd: er bestaan bepaalde erfelijke eenheden, factoren of genen, die voor het totstandkomen van een bepaalde eigenschap verantwoordelijk gesteld mogen worden.
En de localisatie dier genen? De vijftig jaar, die sedert Weismanns hypothese verloopen zijn, hebben ons de erfelijkheidsleer als een zelfstandige, rijkbewerkte wetenschap gebracht. Het door Mendel gegrondveste experiment brak zich baan: gegeven na gegeven leverde elken dag een nieuwe bevestiging van zijn veronderstelling. Het door de Duitsche onderzoekers van 1880 verrichte werk over den microscopischen bouw van cel, celkern en chromosomen werd verdiept en verfijnd en met steeds grooter duidelijkheid kwam het belang van de chromosomen als dragers der erfelijke eigenschappen naar voren. Omstreeks 1902 en 1903 werden door Sutton, Correns en Hugo de Vries, zij het nog
| |
| |
aarzelend, vermoedens geuit, die uitspraken, dat de afzonderlijke factoren, die in grooten getale in elk individu aanwezig zijn, en die elk voor zich een bepaalde eigenschap te voorschijn roepen, in groepen over de verschillende chromosomen verdeeld zouden moeten zijn. In 1904 gaf Boveri de profetische uitspraak: ‘Wanneer een bastaardeering op talrijke eigenschappen betrekking heeft en bij voortgezette teelt blijkt, dat het aantal combinaties, waarin deze eigenschappen met elkaar kunnen samenkomen, grooter is, dan het aantal combinatiemogelijkeden der aanwezige chromosomen, dan leidt dit tot de conclusie, dat de in één chromosoom gelocaliseerde eigenschappen zich bij de reductiedeeling onafhankelijk van elkaar over de beide dochtercellen kunnen verdeelen, wat dus wijst op een uitwisseling van gedeelten der homologe chromosomen’ (d.w.z. de chromosomen, die een paar vormen en bij de reductiedeeling over de beide dochterkernen verdeeld worden) (1904, p. 118).
Profetisch zou deze uitspraak zeker blijken te zijn, want eerst in 1910 begonnen bewijzen geleverd te worden voor de stelling, dat inderdaad de genen in groepen in de chromosomen gelegen zijn. Het was Th.H. Morgan, die met zijn medewerkers de hypothesen van Weismann en anderen tot een localisatietheorie heeft uitgewerkt en ons het experimenteele bewijs heeft verschaft. Merkwaardig genoeg, uit hij zich in Augustus van dat jaar 1910 nog sceptisch: ‘Since the number of chromosomes is relatively small and the characters of the individual are very numerous, it follows on the theory that many characters must be contained in the same chromosome. Do the facts conform to this requisite of the hypotheses? It seems to me that they do not. A few characters, it is true, seem to go together but their number is small, and it is by no means evident that their combination is due to a common chromosome. It is true that in no one species do we know much concerning the behaviour of many characters, but so far as we do know them there is no evidence, that they Mendelize in groups commensurate with the number of chromosomes... But even admitting this possible way of eluding the objection, the other point raised above concerning the absence of groupings of characters in Mendelian inheritance seems a fatal objection to the chromosome theory, so long as that theory attempts to locate each character in a special chromosome’ (1910, p. 467-468).
Inderdaad een merkwaardige sceptische uitlating, indien men in aanmerking neemt, dat juist in dat jaar 1910 in Morgans eigen laboratorium een reeks proeven zou worden ondernomen, die
| |
| |
een afdoende bevestiging van de chromosomenhypothese zouden leveren en haar tot den rang van een goedgefundeerde theorie zouden verheffen. Immers dit onderzoek bij een daartoe bij uitstek geschikt organisme, de bananenvlieg Drosophila, leidde tot de conclusie, dat alle (ongeveer 500) onderzochte eigenschappen, die elk voor zich door een enkel gen beheerscht worden, tot vier groepen behooren, en dat het aantal chromosomen in de voortplantingscellen van dit dier óók vier bedraagt. Dit uitgangspunt en een onafzienbare reeks andere gegevens hebben het onweerlegbare bewijs gebracht, dat de (naar schatting wel eenige duizenden) factoren, die in dit organisme aanwezig zijn, als vier groepen elk in een der vier chromosomen gelegen zijn, dat de afzonderlijke genen in die chromosomen op een rij zijn gerangschikt en daar door een of andere stoffelijke eenheid zijn vertegenwoordigd. Men kan zich het beeld het beste misschien zoo voorstellen: de beide chromosomen, die een paar vormen, zijn de beide huizenrijen in een straat; in ieder huis wonen een of enkele genen, en van elk gen, dat voor een bepaalde eigenschap verantwoordelijk is, kan de straatnaam en het huisnummer worden vastgesteld; het adresboek der genen kan worden opgemaakt.
Intusschen blijft dit chromosoom met zijn genen-inhoud ons nog genoeg moeilijkheden bereiden, die aan de verklaring van de physisch-chemische problemen van chromosomenleven en -functie voorloopig nog in den weg staan. Het heeft allen schijn, alsof het chromosoom een ingewikkeld chemisch systeem van eiwitachtigen aard vormt, waarin dan de genen als speciale op bijzondere plaatsen gelegen zijketens aan het geheel zouden zijn aangekoppeld. Vragen blijven er nog talrijke over, antwoorden zijn nog slechts in klein aantal met eenige waarschijnlijkheid te geven. Maar zeker is wel, dat Weismanns oorspronkelijke en voor zijn tijd zeer gedurfde hypothese weliswaar niet in uitwerking, maar wel in beginsel door het moderne onderzoek bevestigd is, en wij tegenwoordig, misschien onbewust, op zijn werk en zijn gedachtengang voortbouwen.
M.J. Sirks
| |
| |
| |
Literatuur
| Boveri, Th., 1904. Ergebnisse über die Konstitution der chromatischen Substanz des Zellkerns (Jena, Fischer, 1904: 130 pp.). |
| Darwin, Ch., 1868. The variation of animals and plants under domestication (London, Murray, 1868: 411 + 486 pp.). |
| Galton, F., 1871. Experiments in pangenesis by breeding from rabbits of a pure variety, into whose circulation blood taken from other varieties had previously been largely transfused (Proc. Roy. Soc. 19: 393-410). |
| Galton, F., 1875. A theory of heredity (Journ. Anthrop. Inst. London, 5: 329-348). |
| Galton, F., 1877. Typical laws of heredity (Nature, 1877: 492-495, 512-514, 432-533). |
| Galton, F., 1889. Natural inheritance (London, Macmillan, 1889: 259 pp.). |
| Galton, F., 1897. A new law of heredity (Nature, 56: 235-237). |
| Galton, F., 1897. The average contribution of each several ancestor to the total heritage of the offspring (Proc. Roy. Soc., London, 61: 401-413). |
| Galton, F., 1898. A diagram of heredity (illustrating the ‘Ancestral Law’) (Nature, 57: 293). |
| Geurts, P.M.M., 1941. De erfelijkheid in de oudere Grieksche wetenschap (Nijmegen-Utrecht, Dekker & van de Vegt, 1941: 216 pp.). |
| Haeckel, E., 1866. Generelle Morphologie der Organismen (Berlin, Reimer, 1866, I: 574 pp., II: 462 pp.). |
| Koelliker, A., 1885. Die Bedeutung des Zellkerns für die Vorgänge der Vererbung (Zeitschr. f. wiss. Zoolog., 42: 1-46). |
| Morgan, Th.H., 1910. Chromosomes and heredity (Amer. Natur., 44: 449-496). |
| Morgan, Th.H., 1926. The theory of the gene (New Haven, Yale Univ. Press, 1926: 343 pp.). |
| Nägeli, C.W.v., 1884. Mechanisch-physiologische Theorie der Abstammungslehre (München-Leipzig, Oldenbourg, 1884: 822 pp.). |
| Spencer, H., 1864. Principles of biology (London, Williams and Norgate, 2 dln. 1864: 492 + 569 pp.). |
| Strasburger, E., 1884. Neue Untersuchungen über den Befruchtungsvorgang bei den Phanerogamen als Grundlage für eine Theorie der Zeugung (Jena, Fischer, 1884: 176 pp.). |
| Vries, H. de, 1889. Intracellulare Pangenesis (Jena, Fischer, 1889: 212 pp.). |
| Weismann, A., 1882. Ueber die Dauer des Lebens (Jena, Fischer, 1882: 72 pp.). |
| Weismann, A., 1883. Ueber die Vererbung (Jena, Fischer, 1883: 50 pp.). |
| Weismann, A., 1885. Die Continuität des Keimplasmas als Grundlage einer Theorie der Vererbung (Jena, Fischer, 1885: 102 pp.). |
| Weismann, A., 1892. Das Keimplasma. Eine Theorie der Vererbung (Jena, Fischer, 1892: 628 pp.). |
|
|
